 2023, Vol. 35
2023, Vol. 35 



(2: 四川大学水利水电学院, 成都 610065)
(3: 南京水利科学研究院水文水资源与水利工程科学国家重点实验室, 南京 210029)
(2: College of Water Conservancy and Hydropower, Sichuan University, Chengdu 610065, P. R. China)
(3: State Key Laboratory for Hydrology-Water Resources and Hydraulic Engineering, Nanjing Hydraulic Research Institute, Nanjing 210029, P. R. China)
全球约有一半的河流受到大坝建设影响[1]。大坝的建设使河流失去了其天然特性,影响了自然河流水文节律,均化了水温过程,削弱了河道自然洪枯过程[2];同时也改变了生源要素迁移转化过程,从而影响生物地球化学循环[3-4]。水量、水质、栖息地、养分、泥沙输移、水力停留时间等重要因素在水库蓄水后均发生巨大改变,大坝上游被淹没的河段由流动生态系统转变为静止生态系统[5]。被淹没的土地中有机物分解释放养分,提高了富营养化的风险[6-7]。此外,水库拦截作用减少了向下游输送的营养物质,影响营养盐的转化过程,从而改变了水体营养盐的结构特征,造成营养盐失衡[8]。
浮游植物作为水生态系统的初级生产者[9],在物质循环过程中起到了重要介导作用。作为简单的单细胞生物群体,由于种群世代周期短,能够在水库蓄水后快速建立新的群落,被广泛用于评估大型水库对水生态的影响[10-11]。Rangel等通过主成分分析(PCA)发现水力停留时间和磷浓度是热带地区8座水电站浮游植物变化特征的关键环境要素[6];Rodrigues等基于STATICO分析了热带水库浮游植物功能群丰度的时空分布,认为降雨是影响浮游植物组装过程的重要环境因素[12];Nogueira等相关性分析结果显示:水温升高、水力停留时间增加以及养分的富集均会导致浮游植物大量增殖,引起水库发生富营养化[13];Soares基于冗余分析(RDA)发现影响巴西Funil水库浮游植物生物量空间异质性的关键环境要素是水力停留时间[14]。Zhang等基于广义相加模型(GAM)建立硅藻丰度与驱动因子之间的关系,结果表明温度和硅酸盐是饮用水水库中硅藻季节变化的主要驱动因素[15]。由此可见,不同水文水动力条件,海拔、温度等地理因素及氮磷营养盐等共同影响水库浮游植物群落结构[9, 16-18]。在梯级电站中,水库的物理属性、水体理化指标等环境要素的不同会导致浮游植物群落结构和生物量存在差异,如何识别各环境要素对浮游植物群落结构和生物量变化的影响,确定形成梯级水库浮游植物生物量和群落结构差异的主控因子尚有待深入研究。
澜沧江作为西南地区典型的梯级筑坝河流,由北至南,海拔落差大,上下游气候差异明显,沿程分布着不同物理属性、水体理化性质的大小水库,为研究水库的物理属性、水体理化性质与浮游植物群落间的关系提供了良好案例。本研究选择澜沧江自然河道与各级水库开展了多次调研,对比分析不同水库特征下浮游植物生物量和群落结构的差异,并基于GAM模型识别导致梯级水库浮游植物生物量、群落结构(多样性和丰富度)差异的主控因素,以期为水库建设、运行调控以及梯级筑坝河流的科学规划管理提供理论基础。
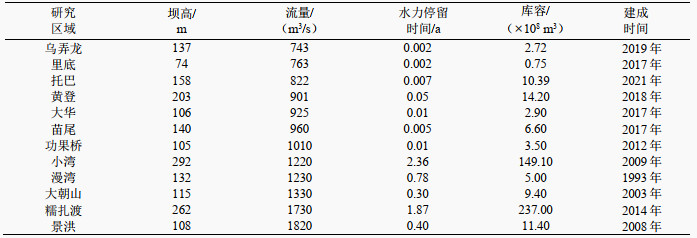
1 材料与方法 1.1 研究区域澜沧江-湄公河是世界第十二大国际河流,发源于唐古拉山北麓的扎曲和昂曲2条河流,于越南胡志明市汇入南海,自北向南全长4880 km[9, 19-20],澜沧江是湄公河的上游[21],流经西藏、青海、云南三省,共53个县[22]。澜沧江上游至下游西双版纳,气温及降雨量呈明显上升趋势,上游属于温带高原气候,年均气温10℃,中游属于亚热带气候,年均气温12~15℃,下游属于亚热带或热带气候,年均气温15~22℃。澜沧江干湿两季分明,丰水期为5—10月,枯水期为11月至次年4月[23]。为开发澜沧江水电能源,已建成乌弄龙、里底、托巴、黄登、大华、苗尾、功果桥、小湾、漫湾、大朝山、糯扎渡、景洪12座水电站,澜沧江梯级水库基本特征参数如附表Ⅰ所示[24]。
1.2 样品采集与测定分别于2016年9月,2018年及2021年的4月和9月进行5次水样和浮游植物样品采集。2016年采集功果桥、小湾、漫湾、大朝山、糯扎渡、景洪的浮游植物样品,2018年在2016年采样点位的基础上增加上游河道、黄登、大华、苗尾、关累5个采样点,2021年在2018年的基础上增加了乌弄龙、里底、托巴3个采样点。研究区及采样点布设如图 1所示。

|
图 1 澜沧江采样点分布 Fig.1 Distribution of sampling sites of Lancang River |
于水面下0.5 m处采集1 L水样,现场加入15 mL鲁哥试剂进行固定,带回实验室内静置48 h后,采用虹吸法去除上清液,将浮游植物样品浓缩至50 mL于定量瓶中保存[9]。浮游植物种类鉴定参考《中国淡水藻类——系统、分类及生态》[25],在10×40倍光学显微镜下镜检及计数。定量计数前充分混合均匀浓缩样品,吸取0.1 mL样品放置于计数框中,观察100个视野,每个样品计数2次,若计数结果相差15 % 以上,再进行第3次计数,取平均值计算细胞丰度,依据形态相似几何学方法计算体积,按照浮游植物的体积公式换算成浮游植物生物量[9, 26]。
野外采用便携式YSI多参数检测仪测定水温(WT)、溶解氧(DO)及pH值,常规水质理化指标总氮(TN)、氨氮(NH3-N)、硝态氮(NO3--N)、亚硝态氮(NO2--N)、总磷(TP)、正磷酸盐(PO43--P)的测定参照《水和废水监测分析方法》(第四版)进行测定[27]。
1.3 数据处理物种多样性指数和丰富度指数(D)被用于分析浮游植物群落多样性及数量动态特点。多样性指数的计算采用Shannon-Wiener指数(H′)进行[9],D采用Margalef的计算公式[28],分别为:
| H′=−s∑i=1(niN)log2(niN) | (1) |
| D=(S−1)/log2N | (2) |
式中,ni为第i种浮游植物的总个体数,N为浮游植物所有种类的总个体数,S为浮游植物样品中种类总数[9, 28]。
广义相加模型(generalized additive model,GAM)是广义线性模型(generalized linear model,GLM)非参数化拓展的分析模型[29-30]。GAM模型可用于处理复杂的响应变量与预测变量之间的非线性、非单调的函数关系[31-32],通过对不同形式函数进行加和,找出其中规律,很好地反映数据特性[33-34]。常用的矩阵排序分析方法是一种用于分析生物种群空间分布及其动态变化机制的线性分析方法,无法解释响应变量与环境要素之间的非线性关系,而GAM模型可用于建立浮游植物长期动态变化趋势分析,在湖泊水库中也常被用于量化相关环境变量的相对贡献,综合考虑变量之间的关系[15, 35-37]。基于GAM模型分别建立丰水期浮游植物群落结构及生物量与关键环境要素的关系,表达式为:
| g(y)=s0+s1(x1)+⋯+sm(xm)+φ | (3) |
式中,g(y)为因变量的连接函数;s(x)为各自变量的平滑函数;φ为随机残差项。
GAM模型采用R 3.6.2软件进行运算,数据分析通过R中“mgcv”工具包实现[38]。GAM模型分析步骤:(1)运用SPSS软件对澜沧江沿程14个采样点浮游植物多样性指数、丰富度指数与环境因子、浮游植物生物量与各环境因子进行Pearson相关性分析,确定主要解释变量;依据相关系数的大小,进行共曲线性判断,若两个解释变量间存在共曲线性,则需剔除其中的一个变量;(2)基于Pearson相关性分析结果,确定GAM模型的非线性拟合方程。自由度(edf)、显著性P值、解释率(deviance explained,D-E)、拟合图像被用于表征GAM模型统计结果。其中edf用于判断因变量与各自变量之间是否属于线性关系,当edf=1表明变量间是线性关系,edf值越大非线性影响能力越强;P值代表统计结果的显著性,其中“***”代表P < 0.001,说明统计结果极显著,其可信度越高;“*”代表P < 0.05,统计结果显著相关[39];D-E用于表征对因变量总体变化的解释率[34],GAM模型拟合的F值代表环境因子重要程度,F值越大,环境因子的重要性越大,可用于量化解释变量对响应变量的贡献程度[40]。
1.4 对比分析影响梯级水库浮游植物群落结构特征的参数主要包括水库物理属性和水体理化指标,其中物理属性主要包括水力停留时间(HRT)、WT、距上游河道的距离等,水体理化指标主要包括氮磷营养盐、DO等指标。为揭示各环境要素对浮游植物生物量和群落结构的影响,分别以HRT和地理距离作为主要变量,定义HRT>1.0 a的水库为大库,HRT < 1.0 a的水库为小库,分别选取上游河道与典型水库进行组合,对比分析自然河道与典型水库间浮游植物群落结构特征的差异。
以HRT作为变量进行距离邻近点位的对比分析:选择上游河道(流速较大,其HRT接近于0)与相邻的首级水库进行对比,其中2018年首级水库为黄登(HRT=0.05 a)、2021年首级水库为乌弄龙(HRT=0.002 a),在本文中统称为邻近水库;选择HRT最大的小湾水库(HRT=2.36 a)与其邻近的功果桥水库(HRT=0.01 a)进行对比分析。
以地理距离作为变量进行相似HRT点位的对比分析:选择小湾(HRT=2.36 a)与糯扎渡(HRT=1.87 a)作为大库的代表水库。糯扎渡位于澜沧江下游,其被视为下游大库,小湾位于糯扎渡上游,被视为上游大库;选择黄登(HRT=0.05 a)和景洪(HRT=0.40 a)作为小库的代表,黄登位于澜沧江上游,其被视为上游小库,景洪为最后一级水库,其被视为下游小库。
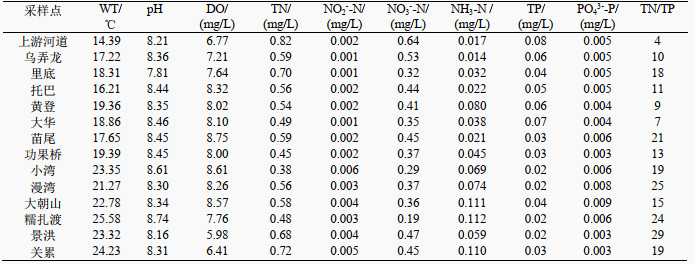
2 结果 2.1 澜沧江各水库水体理化因子分析附表Ⅱ为澜沧江丰水期水体理化指标,澜沧江上游自然河道段主要以冰山融雪为主,WT较低,为14.39℃,远低于各级水库的WT,沿程各水库的WT随着海拔降低呈升高的趋势,至下游河道段(关累),其WT为24.23℃。澜沧江丰水期各采样点的pH值的变化范围为7.81~8.74;DO浓度的变化范围5.98~8.75 mg/L,整体处于地表水Ⅰ类水质标准;TN浓度介于0.38~0.82 mg/L之间,属于地表水Ⅲ类水质标准;TP浓度变化范围为0.02~0.09 mg/L,属于地表水Ⅳ类水质标准。总体而言,澜沧江水质状况整体较好,属于贫营养河流。
2.2 澜沧江浮游植物群落沿程分布特征澜沧江丰水期、枯水期浮游植物生物量沿程变化如图 2a、2b。丰水期澜沧江浮游植物生物量显著高于枯水期,丰水期、枯水期浮游植物生物量变化范围分别为3.93~29.28和1.28~23.32 mg/L。其中小湾水库浮游植物生物量最高,糯扎渡次之,均以绿藻和蓝藻为主。丰水期糯扎渡浮游植物的生物量为19.96 mg/L,枯水期糯扎渡浮游植物的生物量为12.42 mg/L,其中蓝藻门的生物量分别占浮游植物总生物量的36.57 % 和55.39 %。

|
图 2 澜沧江丰水期、枯水期浮游植物生物量及相对丰度沿程变化 Fig.2 Variation of phytoplankton biomass and relative abundance during wet and dry seasons in the Lancang River |
丰水期、枯水期澜沧江浮游植物物种组成相对丰度变化如图 2c、2d。不论丰水期还是枯水期,浮游植物物种组成均以硅藻门、蓝藻门、绿藻门为主。上游河道及乌弄龙、里底、托巴等上游水库浮游植物以硅藻门为主,相对丰度约为80 %。中下游各级水库硅藻门相对丰度明显减少,浮游植物主要以蓝藻门、绿藻门为主。枯水期上游河道、乌弄龙水库浮游植物的物种组成均为硅藻门,无其他门类,而里底、托巴库区除硅藻门外还有部分蓝藻门及绿藻门。黄登、大华硅藻门浮游植物相对丰度约为60 %,苗尾、功果桥、小湾、漫湾、大朝山、糯扎渡、景洪、关累(下游河道)浮游植物物种组成以蓝藻门、绿藻门为主,硅藻门相对丰度减少。
2.3 丰水期浮游植物群落结构特征对比分析上游河道与河道邻近水库、小湾与功果桥对比分析结果如图 3。结果表明:澜沧江上游河道浮游植物的生物量显著低于邻近水库,其生物量平均值分别为3.93、6.72 mg/L;功果桥浮游植物生物量显著低于小湾水库,功果桥生物量平均值为6.75 mg/L,小湾浮游植物生物量为29.29 mg/L。

|
图 3 澜沧江丰水期浮游植物生物量及相对丰度对比分析 (虚线代表分隔线,上游河道vs邻近水库;功果桥vs小湾) Fig.3 Comparative analysis of phytoplankton biomass and relative abundance during wet season in the Lancang River (The dotted line represents the separation line, upstream river vs adjacent reservoir and Gongguoqiao Reservoir vs Xiaowan Reservoir) |
澜沧江丰水期上游河道、邻近水库、小湾及功果桥水库浮游植物相对丰度如图 3b,物种组成以硅藻门、蓝藻门、绿藻门为主,上游河道、邻近水库60 % 以上的浮游植物属于硅藻门,蓝藻门和绿藻门相对丰度较低。功果桥、小湾水库硅藻相对丰度降低,浮游植物物种组成以蓝藻门、绿藻门为主,其相对丰度约为50 %。
上游水库与下游水库之间浮游植物生物量的差异见图 4a,小湾浮游植物生物量高于糯扎渡,糯扎渡浮游植物生物量为17.39 mg/L,上游黄登浮游植物的生物量显著低于下游景洪,黄登浮游植物生物量为7.71 mg/L,景洪浮游植物生物量为14.40 mg/L。

|
图 4 澜沧江丰水期浮游植物生物量(a)及相对丰度(b)对比分析 Fig.4 Comparative analysis of phytoplankton biomass(a)and relative abundance(b) during wet season in the Lancang River |
对比上游小湾、黄登与下游糯扎渡、景洪浮游植物相对丰度(图 4b),结果表明:小湾、糯扎渡库区浮游植物的物种组成以蓝绿藻为主,其中硅藻门相对丰度低于20 %,下游糯扎渡浮游植物物种组成较为丰富;黄登和景洪浮游植物物种组成同样以蓝绿藻门为主,但黄登库区浮游植物硅藻门占比约1/3,明显高于下游景洪库区硅藻的相对丰度。
2.4 丰水期浮游植物群落结构与环境因子GAM模型分析将各级水库物理属性:HRT、WT、距上游河道距离,水库水体理化指标:DO、pH、TN、TP、NH3-N、NO3--N、NO2--N、PO43--P、TN/TP共12项环境要素作为解释变量,浮游植物群落结构(多样性指数、丰富度指数)、生物量分别作为响应变量,建立浮游植物与环境要素的GAM模型。
浮游植物生物量与环境要素GAM模型拟合结果如图 5a~c,图 5i所示,结果表明生物量与HRT相关性极显著(P < 0.001),其次是TN/TP、NO3--N(P < 0.05),与其他环境因子相关性不显著(P> 0.05)。在GAM模型中逐步加入HRT、TN/TP、NO3--N,模型的累积解释率为98.9 %。NO3--N的估计自由度为1,表明NO3--N与浮游植物生物量间是线性关系,而HRT、TN/TP估计自由度分别为2.25和2.85,HRT、TN/TP与生物量间是非线性关系,3个变量的F值排序结果为HRT>TN/TP>NO3--N,其中HRT的值最大,为154.83,说明浮游植物生物量主要受HRT影响。

|
图 5 澜沧江丰水期浮游植物群落结构的GAM模型分析结果 (横坐标为解释变量的实测值;纵坐标代表解释变量对响应变量的拟合值,扩号中的数值则代表估计自由度) Fig.5 Analysis results of GAM model of phytoplankton community structure during wet season in the Lancang River (The abscissa is the measured value of the explanatory variable; the ordinary represents the fitted value of the response variable, the value in the extension represents the estimated degree of freedom) |
基于GAM模型拟合浮游植物群落结构多样性指数、丰富度指数与环境变量间的关系,结果表明多样性指数与HRT、WT、TP的相关性显著(P < 0.05),而与DO、pH、TN、NH3-N、NO3--N、NO2--N、PO43--P的相关性不显著(P > 0.05)。在GAM模型中逐步加入HRT、WT、TP,拟合结果如图 5d~f和图 5i所示,模型的累积解释率为94.4 %,模拟结果较好。HRT、WT、TP的估计自由度分别为2.76、2.23和2.98,其值均大于1,说明HRT、WT、TP与多样性指数呈非线性关系。3个变量的GAM拟合F值排序结果为HRT > TP > WT,表明这些变量对浮游植物的多样性均存在不同程度的影响,其中HRT对浮游植物多样性的影响最大。
图 5g~i为浮游植物丰富度指数与各环境要素GAM模型拟合结果,丰富度指数与NH3-N、TP呈显著相关(P < 0.05),而与其他环境要素相关性不显著(P> 0.05),模型的累积解释率为76.7 %,其中NH3-N的edf=1,表明NH3-N与丰富度指数之间呈线性关系(图 5h),而TP估计自由度为2.87,表明TP与丰富度指数之间呈非线性关系,TP浓度介于0.05~0.10 mg/L之间时,丰富度指数与TP浓度呈负相关关系,当TP浓度介于0.125~0.20 mg/L之间时,丰富度指数与TP浓度增加呈上升趋势(图 5g)。
3 讨论 3.1 澜沧江梯级水库浮游植物生物量时空分布特征及其形成机制澜沧江丰水期浮游植物生物量显著高于枯水期(图 2a、2b),一方面原因是丰水期降雨充沛,土壤中的营养物质通过地表径流的方式被带入库区,为浮游植物生长提供丰富的营养物质;另一方面澜沧江丰水期、枯水期监测的水温平均值分别为20.13、16.78℃,丰水期的水温高于枯水期,更适合浮游植物生长繁殖。小湾和糯扎渡水库的浮游植物生物量均显著高于其他梯级水库,因为小湾和糯扎渡水库HRT最长[24]。HRT是水生态系统中营养物质负荷的关键环境要素,高流速降低了营养物质的可获得性,影响浮游植物的生长[6, 41]。相关研究表明,随HRT增加,水库中营养物质富集,浮游植物丰度也随之增加[10, 13, 42]。
澜沧江上游河道浮游植物生物量显著低于河道邻近水库(黄登、乌弄龙),小湾库区的浮游植物生物量显著高于功果桥(图 3a),在水库建设后,水体流速和紊动强度降低[43],HRT增加、浮游植物对于氮、磷营养物质生物利用度随之提高,促进了浮游植物生长繁殖[42]。上下游大、小水库对比分析结果显示(图 4a),小湾浮游植物的生物量高于糯扎渡,一方面是因为小湾HRT比下游糯扎渡长,另一方面是因为小湾水库位于糯扎渡的上游,其对营养物质存在拦截作用,在一定程度上影响了糯扎渡水库中浮游植物对营养物质的摄取[42]。黄登浮游植物生物量低于下游景洪,原因在于景洪的HRT较长[24],且景洪位于社会经济较为发达的市区,受人类活动扰动强烈,丰水期降雨量增加,大量氮磷营养物质通过地表径流等方式汇入水库,导致营养盐浓度增加;相关研究表明25℃是浮游植物生长繁殖的最佳条件[44],景洪属于亚热带或热带气候,其平均气温高于上游黄登,丰水期的水温维持在24℃左右。较长的HRT、频繁的人类活动、适宜的水温是景洪浮游植物生物量高于黄登的主要原因。
浮游植物生物量GAM拟合结果进一步显示:HRT和营养盐(TN/TP、NO3--N)是影响沿程浮游植物生物量变化的关键环境要素(图 5a~c),其中HRT非线性影响能力最强,TN/TP、NO3--N的影响能力相对较弱,与TN/TP呈非线性关系,与NO3--N呈线性相关。TN/TP是影响浮游植物生物量变化的潜在限制环境要素,Bergström研究认为TN/TP值低于10时,主要表现为氮限制,当TN/TP值高于10时,表现为磷限制[45],苗尾水库至下游河道关累,TN/TP值均高于10(附表Ⅱ),下游各水库处于磷限制的状态。NO3--N是被浮游植物高度吸收利用的氮形态,显著促进了浮游植物生长[46]。
3.2 澜沧江梯级水库浮游植物群落结构时空分布特征及其形成机制对于人工调蓄的水库而言,水库的物理属性、水体理化因子等生境特征共同影响浮游植物群落演替[47]。浮游植物群落结构的组成对食物网动态具有重要生态意义,其变动会影响浮游植物多样性,进而影响水域生态系统功能[48]。丰富度指数可用于表征浮游植物群落的生境物种数目[28],多样性指数可用于反映浮游植物群落结构与食物链上下游生物的营养关系和对生态环境的影响,如:捕食、竞争和演替[49],更多样化的群落能够维持稳定的生态系统功能,如提供生产力、养分保留等[48]。WT、营养盐、HRT共同驱动浮游植物群落结构(多样性、丰富度)变化。
丰水期、枯水期澜沧江上游、下游水库浮游植物物种组成存在明显的空间差异,澜沧江自上游至下游存在1350 m海拔落差,造成上下游的水体存在温差。枯水期(冬季)因上游来水温度较低导致上下游水温差异较大,而丰水期(夏季)上下游温度均有升高且差异较小;高温可提高浮游植物的代谢活性,进而促进了其生物量的升高[50]。雨水对岸边带的冲刷以及抬升的水位对岸边带的淹没,将土壤有机质带入水体中,为浮游植物生长提供营养物质,促进了浮游植物多样性的升高。
丰水期上游至下游(图 4b)浮游植物群落结构呈现硅藻—绿藻—蓝绿藻的变化特征,其群落演变是上下游的气候条件变化所造成的。澜沧江流域由北向南跨越多个气候带,上游西藏属于高原温带气候,下游属于亚热带或热带气候,气温由北向南递增[51]。对比上下游小库,黄登水库的硅藻门相对丰度明显高于景洪,因为硅藻在低温条件下具有极强的耐受性,它可通过调节生理代谢适应低温环境[52]。随着下游WT的升高,蓝藻生长被促进,硅藻的生长受到了抑制。因为蓝藻对高温具有更强的环境适应能力[53],较高的温度可促进其光合作用,延长蓝藻的生长时间[54],WT与多样性指数GAM模型拟合结果(图 5e)显示二者具有非线性关系,合适的温度促进浮游植物种类增加,但是当温度达一定范围,能够快速适应环境的物种大量繁殖反而降低浮游植物多样性,导致生态结构逐渐趋于简单化,影响水生态系统稳定性[54]。
相关研究结果表明营养物质通过各种形式(如有机态和无机态、颗粒态和溶解态、反应性和非反应性)进入水库,库内的生物地球化学循环会改变营养物质的形式和比例,从而影响下游水库生物多样性和生态系统功能,如硅藻门逐渐被蓝藻门、绿藻门所替代[8]。澜沧江水库水体的营养盐浓度总体偏低,营养盐(主要是TP)是影响浮游植物群落结构的关键生境要素,浮游植物多样性指数与TP存在非线性关系(图 5f),当TP浓度介于0~0.035 mg/L时,多样性指数与TP浓度呈现较为显著的正相关关系,当TP浓度进一步增加时,多样性指数反而会随着TP浓度的增加而有所降低。浮游植物丰富度指数与TP存在非线性关系,由于磷元素参与了浮游植物基本生长过程,如光合作用、能量储存以及细胞分裂等[29],需要多样化的浮游植物群落结构维持稳定的生态系统功能[55]。GAM模型拟合结果显示丰富度指数与NH3-N浓度呈线性相关(图 5h),NH3-N是能够被浮游植物直接吸收利用的氮形态[46],NH3-N浓度增加促进了浮游植物丰富度。
上游河道硅藻门相对丰度高于邻近水库,小湾硅藻门相对丰度低于功果桥(图 3b),因为河道与水库、大库与小库的水动力条件不同,上游河道、功果桥HRT显著低于邻近水库及小湾[24],硅藻适合在流速较快、紊动水体环境下生长繁殖[9, 56],因为自身比重较大,较快的流速能够协助硅藻在水体中悬浮[9, 57]。而蓝藻和绿藻更适合在静止、弱紊动水体中生长[58]。因此,澜沧江浮游植物群落结构由上游硅藻演替为下游的蓝藻、绿藻。HRT是营养物质负荷重要调节因素,通过影响浮游植物对营养物质的获取,间接影响了浮游植物多样性,因此,浮游植物多样性先增加后缓慢降低(图 5d)。
4 结论澜沧江梯级水库丰水期浮游植物生物量显著高于枯水期,无论是枯水期还是丰水期,上游水库以硅藻门为主,中下游水库以绿藻门、蓝藻门为主。WT是导致丰水期与枯水期浮游植物生物量差异的主要环境因子,HRT、营养盐是导致上下游浮游植物生物量差异的主要环境因子;WT、营养盐是影响浮游植物群落结构沿程变化的关键因子,HRT则进一步强化了浮游植物群落结构的沿程变化。
5 附录附表Ⅰ和Ⅱ见电子版(DOI: 10.18307/2023.0211)。
| 附表Ⅰ 澜沧江梯级水库基本特征 Appendix Ⅰ Basic characteristics of Lancang River cascade reservoirs |
| 附表Ⅱ 澜沧江丰水期水体理化性质 Appendix Ⅱ Physicochemical characteristics of the water during wet season in Lancang River |
| [1] |
Grill G, Lehner B, Thieme M et al. Mapping the world's free-flowing rivers. Nature, 2019, 569(7755): 215-221. DOI:10.1038/s41586-019-1111-9 |
| [2] |
Wang C. Research conception of ecological protection and restoration of high dams and large reservoirs construction and hydropower cascade development in southwestern China. Advanced Engineering Sciences, 2017, 49(1): 19-26. [王超. "西南水电高坝大库梯级开发的生态保护与恢复"研究构想. 工程科学与技术, 2017, 49(1): 19-26. DOI:10.15961/j.jsuese.2017.01.003] |
| [3] |
Maavara T, Parsons CT, Ridenour C et al. Global phosphorus retention by river damming. Proceedings of the National Academy of Sciences, 2015, 112(51): 15603-15608. DOI:10.1073/pnas.1511797112 |
| [4] |
Poff NL, Richter BD, Arthington AH et al. The ecological limits of hydrologic alteration (ELOHA): A new framework for developing regional environmental flow standards. Freshwater Biology, 2010, 55(1): 147-170. DOI:10.1111/j.1365-2427.2009.02204.x |
| [5] |
Fan H, He DM, Wang HL. Environmental consequences of damming the mainstream Lancang-Mekong River: A review. Earth-Science Reviews, 2015, 146: 77-91. DOI:10.1016/j.earscirev.2015.03.007 |
| [6] |
Rangel LM, Silva LHS, Rosa P et al. Phytoplankton biomass is mainly controlled by hydrology and phosphorus concentrations in tropical hydroelectric reservoirs. Hydrobiologia, 2012, 693(1): 13-28. DOI:10.1007/s10750-012-1083-3 |
| [7] |
Kalff J. Limnology: inland water ecosystems. Prentice-Hall, Upper Saddle River, New Jersey, 2002.
|
| [8] |
Middelburg JJ. Are nutrients retained by river damming?. National Science Review, 2020, 7(9): 1458. DOI:10.1093/nsr/nwaa073 |
| [9] |
Zhang Q, Chen YC, Lin YQ et al. Spatial distribution pattern of phytoplankton community during different water periods in Lancang River cascade reservoir. Acta Scientiae Circumstantiae, 2022, 42(12): 392-401. [张琦, 陈宇琛, 林育青等. 不同水期澜沧江梯级水库浮游植物群落结构空间分布特征. 环境科学学报, 2022, 42(12): 392-401. DOI:10.13671/j.hjkxxb.2022.0150] |
| [10] |
Li JP, Dong SK, Liu SL et al. Effects of cascading hydropower dams on the composition, biomass and biological integrity of phytoplankton assemblages in the middle Lancang-Mekong River. Ecological Engineering, 2013, 60: 316-324. DOI:10.1016/j.ecoleng.2013.07.029 |
| [11] |
Tang J, Cao HQ. Structural adaptions of phytoplankton assemblages along two contrasting reservoirs. Biologia, 2019, 74(2): 139-148. DOI:10.2478/s11756-018-0160-0 |
| [12] |
Rodrigues LC, Pivato BM, Vieira LCG et al. Use of phytoplankton functional groups as a model of spatial and temporal patterns in reservoirs: A case study in a reservoir of central Brazil. Hydrobiologia, 2018, 805(1): 147-161. DOI:10.1007/s10750-017-3289-x |
| [13] |
Nogueira MG, Ferrareze M, Moreira ML et al. Phytoplankton assemblages in a reservoir cascade of a large tropical-subtropical river (SE, Brazil). Brazilian Journal of Biology, 2010, 70(3): 781-793. DOI:10.1590/s1519-69842010000400009 |
| [14] |
Soares MCS, Marinho MM, Azevedo SMOF et al. Eutrophication and retention time affecting spatial heterogeneity in a tropical reservoir. Limnologica, 2012, 42(3): 197-203. DOI:10.1016/j.limno.2011.11.002 |
| [15] |
Zhang Y, Peng CR, Wang J et al. Temperature and silicate are significant driving factors for the seasonal shift of dominant diatoms in a drinking water reservoir. Journal of Oceanology and Limnology, 2019, 37(2): 568-579. DOI:10.1007/s00343-019-8040-1 |
| [16] |
Cui GY, Wang BL, Xiao J et al. Water column stability driving the succession of phytoplankton functional groups in Karst hydroelectric reservoirs. Journal of Hydrology, 2021, 592: 125607. DOI:10.1016/j.jhydrol.2020.125607 |
| [17] |
McGregor GB, Fabbro LD. Dominance of Cylindrospermopsis raciborskii (Nostocales, Cyanoprokaryota) in Queensland tropical and subtropical reservoirs: Implications for monitoring and management. Lakes & Reservoirs: Research & Management, 2000, 5(3): 195-205. DOI:10.1046/j.1440-1770.2000.00115.x |
| [18] |
Burford MA, O'donohue MJ. A comparison of phytoplankton community assemblages in artificially and naturally mixed subtropical water reservoirs. Freshwater Biology, 2006, 51(5): 973-982. DOI:10.1111/j.1365-2427.2006.01536.x |
| [19] |
Chen QW, Chen YC, Yang J et al. Bacterial communities in cascade reservoirs along a large river. Limnology and Oceanography, 2021, 66(12): 4363-4374. DOI:10.1002/lno.11967 |
| [20] |
Zhang LL, Zhao ZQ, Zhang W et al. Characteristics of water chemistry and its indication of chemical weathering in Jinshajiang, Lancangjiang and Nujiang drainage basins. Environmental Earth Sciences, 2016, 75(6): 1-18. DOI:10.1007/s12665-015-5115-y |
| [21] |
Liu M, Han GL, Zeng J et al. Effect of cascade reservoirs on geochemical characteristics of rare earth elements in suspended particle matter in Lancangjiang River, Southwest China. Aquatic Sciences, 2022, 84(2): 1-19. DOI:10.1007/s00027-022-00851-w |
| [22] |
Zeng J, Han GL, Zhang ST et al. Potentially toxic elements in cascade dams-influenced river originated from Tibetan Plateau. Environmental Research, 2022, 208: 112716. DOI:10.1016/j.envres.2022.112716 |
| [23] |
Zhang BY, Ding W, Xu B et al. Spatial characteristics of total phosphorus loads from different sources in the Lancang River Basin. Science of the Total Environment, 2020, 722: 137863. DOI:10.1016/j.scitotenv.2020.137863 |
| [24] |
Ma HH, Chen YC, Chen QW et al. Dam cascade unveils sediment methylmercury dynamics in reservoirs. Water Research, 2022, 212: 118059. DOI:10.1016/j.watres.2022.118059 |
| [25] |
胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态. 北京: 科学出版社, 2006.
|
| [26] |
Sun J, Liu DY, Qian SB. Study on phytoplankton biomass Ⅰ. Phytoplankton measurement biomass from cell volume or plasma volume. Acta Oceanologica Sinica, 1999, 21(2): 75-85. [孙军, 刘东艳, 钱树本. 浮游植物生物量研究Ⅰ. 浮游植物生物量细胞体积转化法. 海洋学报, 1999, 21(2): 75-85.] |
| [27] |
Editorial board of water and wastewater monitoring and analysis method, Ministry of Environmental Protection of the People's Republic of China eds. Water and wastewater monitoring and analysis methods: 4th edition. Beijing: China Environmental Science Press, 2002. [国家环境保护局《水和废水监测分析方法》编委会. 水和废水监测分析方法: 第4版. 北京: 中国环境科学出版社, 2002.]
|
| [28] |
Huang FH, Shi HH, Zheng W et al. Phytoplankton diversity distribution and its influence factors in southern waters of Miaodao Archipelago in summer. Marine Environmental Science, 2015, 34(4): 530-535. [黄风洪, 石洪华, 郑伟等. 夏季庙岛群岛南部海域浮游植物多样性分布及其影响因子. 海洋环境科学, 2015, 34(4): 530-535.] |
| [29] |
Zhang JP, Zhi MM, Zhang Y. Combined generalized additive model and random forest to evaluate the influence of environmental factors on phytoplankton biomass in a large eutrophic lake. Ecological Indicators, 2021, 130: 108082. DOI:10.1016/j.ecolind.2021.108082 |
| [30] |
Yi YJ, Cheng X, Zhou J. Research progress in habitat suitability assessment methods. Ecology and Environmental Sciences, 2013, 22(5): 887-893. [易雨君, 程曦, 周静. 栖息地适宜度评价方法研究进展. 生态环境学报, 2013, 22(5): 887-893.] |
| [31] |
Zhao J, Zhang SY, Wang ZH et al. Fish community diversity distribution and its affecting factors based on GAM model. Chinese Journal of Ecology, 2013, 32(12): 3226-3235. [赵静, 章守宇, 汪振华等. 基于GAM模型的鱼类群落多样性分布及影响因素. 生态学杂志, 2013, 32(12): 3226-3235. DOI:10.13292/j.1000-4890.2013.0494] |
| [32] |
Zhu Y, Kang MY. Application of ordination and GLM/GAM in the research of the relationship between plant species and environment. Chinese Journal of Ecology, 2005, 24(7): 807-811. [朱源, 康慕谊. 排序和广义线性模型与广义可加模型在植物种与环境关系研究中的应用. 生态学杂志, 2005, 24(7): 807-811.] |
| [33] |
Morton R, Henderson BL. Estimation of nonlinear trends in water quality: An improved approach using generalized additive models. Water Resources Research, 2008, 44(7): W07420. DOI:10.1029/2007WR006191 |
| [34] |
Zhang ZY, Niu Y, Yu H et al. Relationship of chlorophyll-a content and environmental factors in lake Taihu based on GAM model. Research of Environmental Sciences, 2018, 31(5): 886-892. [张智渊, 牛远, 余辉等. 基于GAM模型的太湖叶绿素a含量与环境因子相关性分析. 环境科学研究, 2018, 31(5): 886-892. DOI:10.13198/j.issn.1001-6929.2018.03.20] |
| [35] |
Lehtinen S, Suikkanen S, Hällfors H et al. Approach for supporting food web assessments with multi-decadal phytoplankton community analyses-case Baltic Sea. Frontiers in Marine Science, 2016, 3: 220. DOI:10.3389/fmars.2016.00220 |
| [36] |
Xiao WP, Liu X, Irwin AJ et al. Warming and eutrophication combine to restructure diatoms and dinoflagellates. Water Research, 2018, 128: 206-216. DOI:10.1016/j.watres.2017.10.051 |
| [37] |
Stefanidis K, Varlas G, Vourka A et al. Delineating the relative contribution of climate related variables to chlorophyll-a and phytoplankton biomass in lakes using the ERA5-Land climate reanalysis data. Water Research, 2021, 196: 117053. DOI:10.1016/j.watres.2021.117053 |
| [38] |
Wood S. Generalized additive models: An introduction with R. Boca Raton: CRC Press, 2006.
|
| [39] |
Yuan WH, Wang H, Xia YB et al. Relationship of chlorophyll-a and water quality factors in Poyang Lake based on GAM model. Ecology and Environmental Sciences, 2021, 30(8): 1716-1723. [袁伟皓, 王华, 夏玉宝等. 基于GAM模型的鄱阳湖叶绿素a与水质因子相关性分析. 生态环境学报, 2021, 30(8): 1716-1723.] |
| [40] |
Yan GH, Yin XY, Wang X et al. Analysis based on GAM model for the relationship between chlorophyll-a concentration and environmental factors in Dongting Lake. China Environmental Science, 2022, 42(1): 313-322. [严广寒, 殷雪妍, 汪星等. 基于GAM模型的洞庭湖叶绿素a浓度与环境因子相关性分析. 中国环境科学, 2022, 42(1): 313-322.] |
| [41] |
Schindler DW. Recent advances in the understanding and management of eutrophication. Limnology and Oceanography, 2006, 51(1part2): 356-363. DOI:10.4319/lo.2006.51.1_part_2.0356 |
| [42] |
Chen QW, Shi WQ, Huisman J et al. Hydropower reservoirs on the upper Mekong River modify nutrient bioavailability downstream. National Science Review, 2020, 7(9): 1449-1457. DOI:10.1093/nsr/nwaa026 |
| [43] |
Syvitski JPM, V r smarty CJ, Kettner AJ et al. Impact of humans on the flux of terrestrial sediment to the global coastal ocean. Science, 2005, 308(5720): 376-380. DOI:10.1126/science.1109454 |
| [44] |
赵颖. 水文、气象因子对藻类生长影响作用的试验研究[学位论文]. 南京: 河海大学, 2006.
|
| [45] |
Bergstr m AK. The use of TN: TP and DIN: TP ratios as indicators for phytoplankton nutrient limitation in oligotrophic lakes affected by N deposition. Aquatic Sciences, 2010, 72(3): 277-281. DOI:10.1007/s00027-010-0132-0 |
| [46] |
Wetzel RG. Limnology: river and lake ecology. San Diego: Academic Press, 2001: 1006.
|
| [47] |
Zhang JL, Zheng BH, Liu DF et al. Succession pattern of phytoplankton of Daning River in the Three Gorges Reservoir and its driving factors. Environmental Science, 2017, 38(2): 535-546. [张佳磊, 郑丙辉, 刘德富等. 三峡水库大宁河支流浮游植物演变过程及其驱动因素. 环境科学, 2017, 38(2): 535-546. DOI:10.13227/j.hjkx.201606137] |
| [48] |
Amorim CA, Moura ADN. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Science of the Total Environment, 2021, 758: 143605. DOI:10.1016/j.scitotenv.2020.143605 |
| [49] |
Stirling G, Wilsey B. Empirical relationships between species richness, evenness, and proportional diversity. The American Naturalist, 2001, 158(3): 286-299. DOI:10.1086/321317 |
| [50] |
Li HF, Lin SJ. Research progresses of phytoplankton ecology in the South China Sea. Journal of Xiamen University: Natural Science, 2019, 58(1): 1-10. [李红飞, 林森杰. 南海浮游植物生态学研究进展. 厦门大学学报: 自然科学版, 2019, 58(1): 1-10.] |
| [51] |
Zhang JH, Feng ZM, Jiang LG et al. Analysis of the correlation between NDVI and climate factors in the lancang river basin. Journal of Natural Resources, 2015, 30(9): 1425-1435. [张景华, 封志明, 姜鲁光等. 澜沧江流域植被NDVI与气候因子的相关性分析. 自然资源学报, 2015, 30(9): 1425-1435.] |
| [52] |
Li J, Yin W, Jia HY et al. Research progress on diatom blooms in the middle and lower Hanjiang River: Review and advances. Journal of Hydroecology, 2020, 41(5): 136-144. [李建, 尹炜, 贾海燕等. 汉江中下游硅藻水华研究进展与展望. 水生态学杂志, 2020, 41(5): 136-144. DOI:10.15928/j.1674-3075.2020.05.016] |
| [53] |
Zhu GW, Shi K, Li W et al. Seasonal forecast method of cyanobacterial bloom intensity in eutrophic Lake Taihu, China. J Lake Sci, 2020, 32(5): 1421-1431. [朱广伟, 施坤, 李未等. 太湖蓝藻水华的年度情势预测方法探讨. 湖泊科学, 2020, 32(5): 1421-1431. DOI:10.18307/2020.0504] |
| [54] |
Rasconi S, Winter K, Kainz MJ. Temperature increase and fluctuation induce phytoplankton biodiversity loss-Evidence from a multi-seasonal mesocosm experiment. Ecology and Evolution, 2017, 7(9): 2936-2946. DOI:10.1002/ece3.2889 |
| [55] |
Xu SJL, Chan SCY, Wong BYK et al. Relationship between phytoplankton community and water parameters in planted fringing mangrove area in South China. Science of the Total Environment, 2022, 817: 152838. DOI:10.1016/j.scitotenv.2021.152838 |
| [56] |
Li FP, Gao Y, Zhang HP et al. Simulation experiment on the effect of flow velocity on phytoplankton growth and composition. J Lake Sci, 2015, 27(1): 44-49. [李飞鹏, 高雅, 张海平等. 流速对浮游藻类生长和种群变化影响的模拟试验. 湖泊科学, 2015, 27(1): 44-49. DOI:10.18307/2015.0106] |
| [57] |
Harris GP. Temporal and spatial scales in phytoplankton ecology: mechanisms, methods, models, and management. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(5): 877-900. DOI:10.1139/f80-117 |
| [58] |
Li G, Lin ML, Chen C et al. Autumn and winter characteristics of phytoplankton community structure and its relationship with environmental factors in the northern Jiangsu section of grand canal. Journal of Hydroecology, 2021, 42(5): 119-126. [李港, 林妙丽, 陈诚等. 京杭运河苏北段秋冬季浮游植物群落结构特征及其与环境因子的关系. 水生态学杂志, 2021, 42(5): 119-126. DOI:10.15928/j.1674-3075.202105250164] |