 2023, Vol. 35
2023, Vol. 35 



(2: 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072)
(3: 青海省生态环境监测中心, 国家环境保护青藏高原生态环境监测与评估重点实验室实验室, 青海省生态环境监测与评估重点实验室, 西宁 810007)
(2: State Key Laboratory of Fresh Water Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, P. R. China)
(3: Qinghai Eco-Environment Monitoring Center, State Environmental Protection Key Laboratory of Tibetan Plateau Eco-Environmental Monitoring and Assessment, Qinghai Key Laboratory of Eco-Environmental Monitoring and Assessment, Xining 810007, P. R. China)
青藏高原位于亚欧大陆中南部,是世界上海拔最高的高原[1],被誉为“亚洲水塔”,不仅是长江和黄河的发源地,还是南亚、东南亚和中南半岛等地区诸多重要河流的发源地[2]。青藏高原面积在1 km2以上的湖泊有1171个,是全球湖泊分布最密集的地区之一。由于高海拔区域低温、强辐射等极端的环境条件,青藏高原区域生态环境简单且脆弱,极易遭受破坏[3]。研究表明,青藏高原气候变暖的速度是全球平均速度的3倍[4],在全球变暖和人类活动的干扰下,青藏高原地区正遭受着冰川消融、水土失衡等多种生态环境问题[5-6]。
高海拔的湖泊生态系统是监测环境变化、人为干扰以及全球气候变化的理想对象,因为这类生态系统对外界环境变化的反应往往更为迅速和敏感[7-8],即使是相对较小的环境扰动也会显著影响这些湖泊的物理化学性质,引起物种组成和生物群落的变化[9]。近年来,高海拔湖泊作为生物多样性重点保护对象,越来越受到人们的重视[7],国际上针对高原湖泊中水生生物群落时空格局、水生生物对气候变化及环境干扰的响应开展了大量的研究[10-11]。而在青藏高原的湖泊中,目前关于水生生物的研究却仍然十分缺乏,阻碍了这些湖泊的有效管理和保护[1, 4]。
浮游植物作为水生态系统中最重要的初级生产者,占据了全球净初级生产力的50 % 以上,它们在水生态系统的物质循环和能量流动中起着至关重要的作用[12-13]。相比其它水生生物,浮游植物生长周期短,对环境变化能作出快速响应。浮游植物群落结构的季节性演替能够表征水环境的状态,同时水环境的时空变化会影响浮游植物的组成与分布[14-15]。在高海拔湖泊中,海拔、盐度和水温变化通常会对浮游植物群落产生较大的影响[16-17]。国内外关于浮游植物群落时空格局及其驱动力也开展了许多研究,但这些研究多集中于平原地区的水体[18-19],而有关高原湖泊的研究还鲜见报道。
青海湖是位于青藏高原的典型咸水湖泊,也是中国最大的内陆咸水湖,处于青藏高原高寒区和黄土高原干旱区的过渡地带。近些年来,在周边社会经济发展特别是旅游业发展及全球气候变化的大背景下,青海湖面临着如水位上升、沙漠化加剧、草场退化、鱼类资源锐减等一系列水生态环境问题[20-21]。目前国内外有关青海湖的研究主要集中在气候变化[22-23]、水位状况[24-25]、植被覆盖[26-27]和鱼类资源[28-29]等领域,有关该区域的浮游植物研究却相对薄弱,现有的少量研究主要集中于种类记录或单个季度的群落调查[30-34],有关浮游植物群落时空尺度上(不同季节、不同区域)的研究十分罕见。
鉴于青海湖高原咸水湖泊的特殊性及浮游植物群落时空格局的未知现状,本文于2020年5月、8月和10月(分别对应春、夏、秋)3个季节对青海湖主湖区20个采样点的浮游植物进行调查,分析青海湖浮游植物群落组成和环境因子的时空特征,并对浮游植物与环境因子之间的关系进行探究,旨在阐明青海湖浮游植物群落结构和时空分布格局,为青海湖的管理、保护以及水生生物资源的可持续发展提供科学依据。
1 材料与方法 1.1 研究区域概况及采样点布置 1.1.1 湖泊概况青海湖(36°32′~37°15′N,99°36′~100°47′E)地处青藏高原东北部,水域面积4625.6 km2,东西长约105 km,南北宽约63 km,湖面平均海拔3294.93 m。青海湖地处高原半干旱高寒气候区,年平均气温在-0.8~1.1℃之间,每年11月至次年3月为冰封期,年平均冰封期为100~129 d。
1.1.2 样点布设于2020年5月(春季)、8月(夏季)、10月(秋季)对青海湖全湖区进行采样调查(其中,8月份由于青海湖风浪影响,实际采样调查的结束时间顺延到了9月初)。依据水深以及青海湖入湖河流输入路线,将青海湖划分为4个区域:湖滨带(N1~N4,水深 < 12 m,距离岸边较近,无河流汇入区域)、进水区(N5~N11,水深 < 12 m,入湖支流汇入区域)、浅水区(N12~N17,水深12~24 m)和深水区(N18~N20,水深> 24 m),共设置20个采样点(图 1)。生物采样与水化学采样同步进行。

|
图 1 青海湖采样点分布 Fig.1 Distribution of sampling sites in Lake Qinghai |
现场使用多参数水质分析仪(HQ40D, Hach Love-land, USA)测定各样点溶解氧(DO)、水温(WT)、pH、盐度(salinity)和氯离子(Cl-)等水体理化指标。在每个采样点采集表层(0.5 m)水样1 L,装于样品瓶中,低温避光保存,24 h内带回实验室,测定高锰酸盐指数(CODMn)、生化需氧量(BOD5)、化学需氧量(COD)、总氮(TN)、氨氮(NH3-N)、硝态氮(NO3--N)和总磷(TP)浓度。水样保存及测定方法均参照国家标准[35]。
1.3 浮游植物样品的采集与鉴定用5 L有机玻璃采水器采集湖泊水体表层(0.5 m)水样,取1 L水样装入广口瓶中,加入15 mL鲁哥氏液固定,沉淀48 h后浓缩至50 mL,充分摇匀后吸取0.1 mL于容积为0.1 mL的计数框中,在光学显微镜(Olympus CX21)下进行计数。浮游植物鉴定参考《中国淡水藻类——系统、分类及生态》[36]和《中国淡水生物图谱》[37],采用细胞体积法计算浮游植物生物量。
1.4 数据处理采用Shannon-Wiener多样性指数(H′)[38]、Simpson多样性指数(D)[39]、Margalef丰富度指数(d)[40]和Pielou均匀度指数(J)[41]表示浮游植物群落多样性,计算公式分别如下:
| H′=−s∑i=1PilnPi | (1) |
| D=1−s∑i=1P2i | (2) |
| d=(S−1)/lnN | (3) |
| J=H′/lnS | (4) |
式中,Pi为第i种藻类个体数(Ni)占总个体数(N)的比例,S为样点物种数。
优势种根据各个物种优势度(Y)值确定,公式为:
| Y=Ni/N×fi | (5) |
式中,fi为第i种类出现的频率。将Y≥0.02的种类为优势种[42]。
使用Kruskal-Wallis检验分析环境变量在不同区域和季节间的差异。使用单因素重复测量方差分析(One-way,repeated-Measures ANOVA)比较湖区4个区域和3个季节浮游植物物种数、丰度、生物量和多样性指数之间是否存在显著差异。如果单因素方差分析的组间差异显著,进行多重比较;如果方差齐,进行HSD (Tukey's honestly significant difference)检验;如果方差不齐,选用Games-Howell检验。
基于浮游植物丰度数据,采用非度量多维尺度(Non-metric multidimensional scaling,NMDS)分析浮游植物群落时空格局,进一步使用置换多元方差分析(Permutational multivariate analysis of variance,PERMANOVA)检验不同区域和季节间植物群落结构是否存在差异。
运用基于距离的冗余分析(distance-based redundancy analysis,dbRDA),探究浮游植物群落结构与环境因子之间的关系。在dbRDA型中,将3次调查的生物群落结构组成作为生物数据源,环境因子参数作为环境数据源。为了优化分析,对生物数据进行log(x+1)转化,对不满足正态分布的环境因子进行log(x+1)转化,为避免存在过高共线性,剔除具有较高相关系数(r > 0.8)的环境因子。用前选法(forward selection,基于最高的adjusted R2)确定对群落分布具有重要且独立作用的关键环境因子进入最终的dbRDA中[43-44]。
Kruskal-Wallis检验和单因素重复测量方差分析在SPSS 26.0中完成,NMDS、PERMANOVA和dbRDA分析在PRIMER & PERMANOVA+6.0中完成。
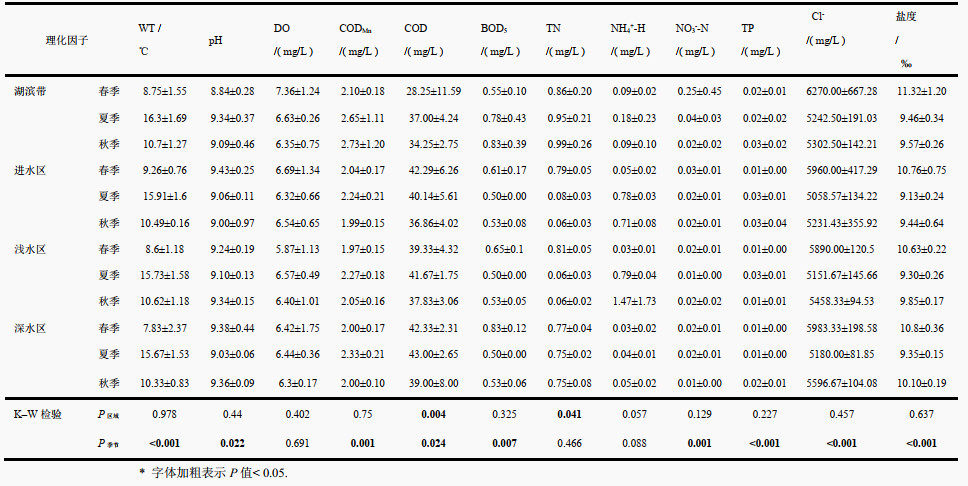
2 结果 2.1 青海湖水环境因子特征对环境因子的K-W检验结果显示,在4个不同的湖区间,湖滨带COD低于其他区域,而TN和NH3-N均高于其他区域,其余理化因子无显著的区域差异。在3个季节间,多数环境因子在不同季节间差异显著。其中,水温、COD和CODMn均在夏季达到最高值,NO3--N、BOD5、Cl-和盐度在春季呈现最高值,pH和TP则在秋季达到最高值(附表Ⅰ)。
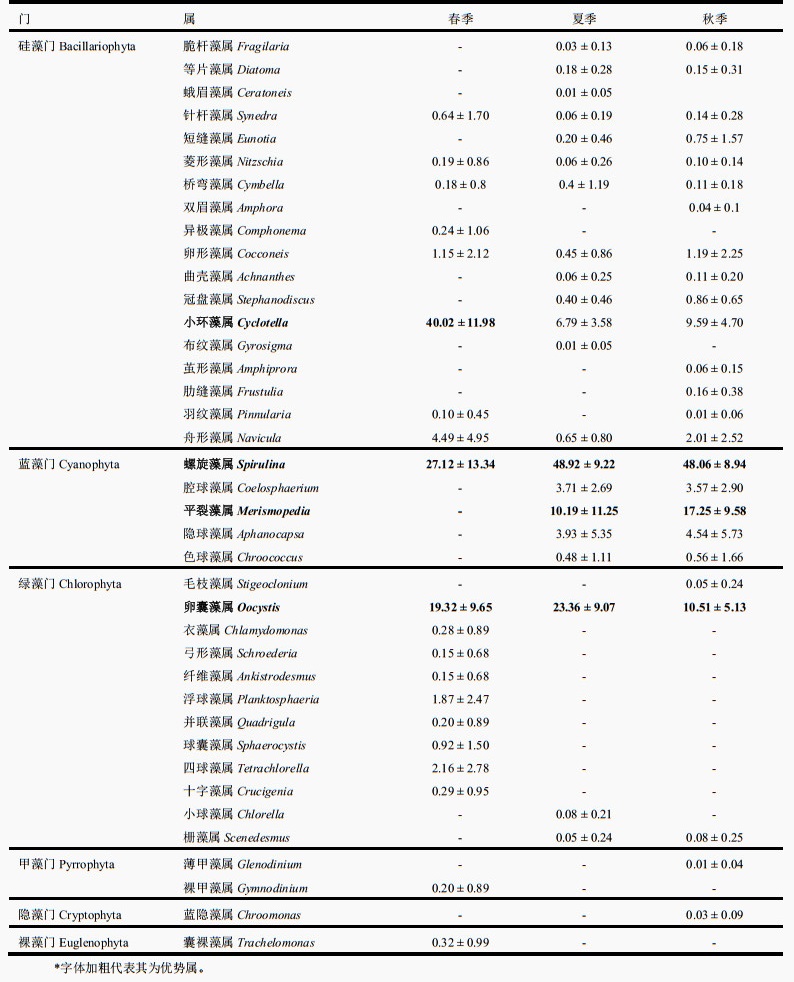
2.2 青海湖浮游植物群落特征3次调查共发现浮游植物6门23科39属65种。其中硅藻门(Bacillariophyta)18属36种(占总物种数的55.38 %);绿藻门(Chlorophyta)12属17种(占26.15 %);蓝藻门(Cyanophyta)5属7种(占10.76 %);甲藻门(Pyrrophyta)2属2种,隐藻门(Cryptophyta)1属2种;裸藻门(Euglenophyta)最少,只有1属1种。春季优势属为小环藻属(Cyclotella)、鞘丝藻属(Lyngbya)和卵囊藻属(Oocystis),夏、秋季优势属则为鞘丝藻属、卵囊藻属和平裂藻属(Merismopedia)(附表Ⅱ)。
3次调查的优势类群均为硅藻门、蓝藻门和绿藻门,但春季与夏、秋季的主要优势类群有一定的差异:春季最具优势类群为硅藻门,其占浮游植物总丰度的47.01 %。夏、秋季最具优势类群则为蓝藻门,占总丰度的比例分别为67.23 % 和73.97 %,而硅藻门比例明显下降(9.28 %,15.34 %);此外,优势种组成在3个季节间也发生了一定变化,尤其春季与夏、秋两季的优势种组成差异较大。春季的优势种类主要为硅藻门的种类,而夏、秋两季的优势种主要为蓝藻门的种类。硅藻门的小环藻(Cyclotella sp.)在春季最占优势,但其在夏、秋两季的优势度明显下降(表 1,图 2)。
| 表 1 青海湖浮游植物优势种及优势度指数 Tab. 1 Dominant species and dominance index of phytoplankton in Lake Qinghai |

|
图 2 青海湖4个区域3个季节优势种群丰度和相对丰度的均值和标准差 Fig.2 Mean value and SD of abundance and relative abundance of dominant groups at 4 regions among 3 seasons in Lake Qinghai |
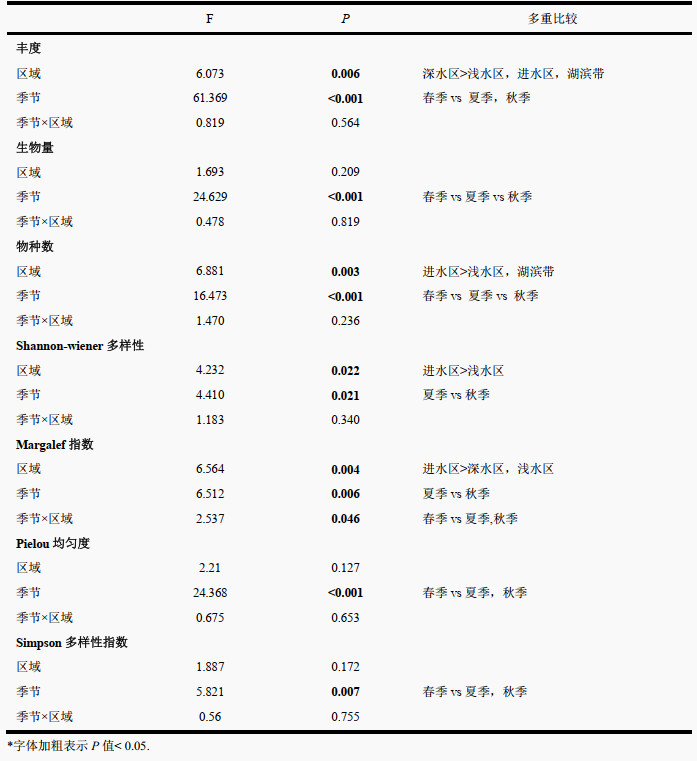
重复测量方差分析结果显示,浮游植物的丰度、物种数、Shannon-Wiener多样性指数和Margalef指数在不同区域和不同季节间有显著差异。区域尺度上,丰度表现为深水区最高(3.44×106 cells/L),浅水区(2.64×106 cells/L)、进水区次之(2.43×106 cells/L),湖滨带最低(2.41×106 cells /L),呈现出深水区向四周辐射递减的趋势;物种数、Shannon-Wiener多样性指数和Maraglef指数则均表现为进水区较高而浅水区较低的趋势,其余指数在各个区域间变化不大(图 3,附表Ⅲ)。

|
图 3 青海湖4个区域3个季节浮游植物丰度、生物量、丰富度和多样性指数 Fig.3 Abundance, biomass, taxa richness, and diversity indexes of phytoplankton at 4 regions among 3 seasons in Lake Qinghai |
3个季节间,春季的丰度和生物量仅为夏、秋季的1/10左右。具体而言,丰度和生物量均在秋季(丰度4.20×106 cells/L,生物量189.14 μg/L)最高,夏季(3.35×106 cells/L,98.56 μg/L)次之,春季(3.55×105 cells/L,9.67 μg/L)最低。物种数和各多样性指数在3个季度间也存在显著差异,但差异不如丰度和生物量差异大:物种数在3个季度间的变化趋势与丰度、生物量变化趋势一致,即春季最低而秋季最高。秋季Margalef指数和Shannon-Wiener多样性指数均高于夏季。Pielou均匀度和Simpson多样性指数在春季较高,夏季和秋季较低(图 3, 附表Ⅲ)。
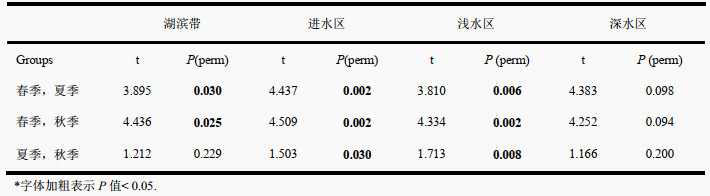
PERMANOVA分析显示,区域(F=1.849,P=0.002)和季节(F=42.114,P=0.001)均是影响浮游植物群落的重要因素,但区域和季节的交互作用对浮游植物群落影响不显著(F=1.132,P=0.207)。进一步的两两对比分析表明,群落结构的区域差异在春季尤为显著,尤其是湖滨带和其他3个区域的群落结构差别较大。在季节尺度上,湖滨带、进水区和浅水区的群落结构在3个季节间均有较大差异。相比之下,深水区较为稳定,在3个季节间均无显著变化(附表Ⅳ、Ⅴ)。群落格局的时空差异在NMDS排序图中也得到了较好的体现,不同季节间群落结构比不同区域间的区分更为明显,尤其是春季与夏、秋两季的群落组成差异较大,而夏、秋两季区别较小,浮游植物群落结构更为接近(表 2、图 4)。
| 表 2 青海湖浮游植物群落的PERMANOVA结果* Tab. 2 Permutational analysis of variance (PERMANOVA) results for the phytoplankton communities in relation to region and season in Lake Qinghai |

|
图 4 青海湖浮游植物多维尺度排序图:区域(a);季节(b) Fig.4 Non-metric multidimensional scaling (NMDS) ordination biplots of phytoplankton samples from Lake Qinghai: region(a); season(b) |
dbRDA分析结果显示,盐度、WT和TP是影响青海湖浮游植物群落时空分布的关键环境因子(P < 0.05),同时,BOD5也接近显著水平(P=0.058)。这4个环境因子共同解释了39.0 % 的群落结构变化(Adj.R2=0.390)(图 5、表 3)。

|
图 5 青海湖浮游植物群落与环境变量的dbRDA排序图:区域(a);季节(b) Fig.5 Distance-based redundancy analysis (dbRDA) of phytoplankton assemblages to environmental variables in Lake Qinghai: region(a); season(b) |
| 表 3 青海湖浮游植物群落结构与环境因子的dbRDA分析结果汇总* Tab. 3 Summary results of distance-based redundancy analyses (dbRDA) between phytoplankton communities and environmental factors in Lake Qinghai |
历史上,青海湖浮游植物调查研究资料较为匮乏,20世纪仅中国科学院兰州地质研究所在1961—1962年[31]及青海省生物研究所在1964—1967年[30]开展过2次大规模的调查,记录了浮游植物属数及丰度。其中,第2次调查的浮游植物属数及丰度较第一次有所增加。进入21世纪以来,杨建新等[32]及姚维志等[33-34]分别在2006年及2006—2010年的夏季在青海湖开展过浮游植物群落调查,与20世纪调查结果相比较,21世纪初的调查发现青海湖浮游植物属的数目有所减少,尤其硅藻门中有9个属在21世纪初的调查中没有发现,这种变化的原因可能在于调查时间不一致(20世纪是四季调查而21世纪的调查只有1个季度)以及湖泊水环境质量的降低[32-34]。王一郎[45]在2019年对青海湖浮游植物的调查结果显示,青海湖浮游植物部分样点丰度达到了107 cells/L,蓝藻门已成为青海湖浮游植物的绝对优势类群。本研究的结果与王一郎[45]调查结果较为相似,与历史上的调查相比,主要有两大变化:1)浮游植物的丰度出现了较大幅度的增加(历史调查浮游植物的丰度基本在104~105 cells/L,而本研究中夏秋两季丰度达到了106 cells/L);2)浮游植物的优势类群发生了较大的变化(20世纪与21世纪初的调查中硅藻门均为最优势类群[11-15],而本研究中夏、秋两季蓝藻门已超过硅藻门成为最优势类群)(表 4)。
| 表 4 不同时期青海湖浮游植物调查数据 Tab. 4 The data of phytoplankton in Lake Qinghai in different investigation periods |
青海湖浮游植物优势类群的结构变化与水温上升、营养盐增加和盐度下降有着密不可分的关系。研究显示,1966—2016年间,非冰期的青海湖表层湖水平均温度整体呈逐渐升高的趋势,增加速率为0.50℃/10 a,高于全球范围内167个大型湖泊的表面温度增加速率(0.45℃/10 a)[46]。水温的上升可改变浮游植物细胞新陈代谢的速率,对酶的活性、营养盐吸收速率和细胞分裂周期等造成不同程度的影响,进而造成浮游植物丰度显著上升[13, 15]。由于浮游植物不同的种类对温度有不同的适应性,硅藻更适合在低温环境中生长而蓝藻更适合温度较高的水体中生长[47]。近年来,由于水位上升导致草场淹没以及周边旅游业的发展,外源氮磷污染物通过降水冲刷等途径进入湖泊,青海湖总磷和总氮浓度均有大幅度升高[48-50]。营养盐浓度变化能够影响浮游植物群落结构组成,不同浮游植物对营养盐的吸收和耐受能力也不一致,适宜浓度范围内的营养盐能够极大地促进浮游植物的生长[51]。有研究表明,蓝藻在营养盐较高的水体中易成为优势种群,而且营养盐对蓝藻的生长促进作用甚至有时大于水温增加对蓝藻的促进作用[52-53]。从21世纪初到现在,青海湖总磷含量逐年升高,相比1960s有大幅度的升高,但总氮浓度年际变动不大(0.74~1.1 mg/L)[49]。这表明低浓度的磷可能是限制青海湖浮游植物丰度的主要因素。近年来磷浓度的增加,促进了浮游植物丰度的增加,同时促使蓝藻在浮游植物群落演替中占优势[54]。相比于其他藻类,硅藻门的种类对盐度具有更大的耐受性,故其在高海拔咸水湖泊中往往成为浮游植物群落的绝对优势类群,因为适当的盐度有利于硅藻吸收硅促使其锥状体的形成[55-57]。从1960s到21世纪初,随着水位下降,青海湖盐度逐年上升(12.49 ‰上升至15.34 ‰),随后盐度开始逐渐下降(15.34 ‰下降至10.32 ‰)[49-50],本研究中,夏、秋两季样点的盐度平均值为9.48 ‰ (最低值为8.12 ‰)。有研究显示,在5 ‰ ~10 ‰的盐度范围内,蓝藻的生长速度最快[58],故近年来盐度的显著下降也可能在一定程度上促进了青海湖蓝藻门种类的生长繁殖。
3.2 青海湖浮游植物时空格局本研究分析结果显示,青海湖浮游植物群落丰度、生物量和物种数存在较明显的时空差异。一般情况下,大型湖泊湖心深水区域浮游植物的丰度通常低于近岸区域[59]。但本次调查却发现青海湖深水区的浮游植物平均丰度高于其他区域,且丰度呈现出深水区向周围辐射递减的趋势。这与姚维志等在青海湖调查结果一致[33-34],这种现象可能与青海湖具体环境条件有关。青海湖水域辽阔,常年盛行的西北风和入湖河流的注入是青海湖湖浪和湖流的主要驱动力[60],较大的风浪通常会对水体造成强烈的扰动,尤其是对大个体的浮游植物细胞产生不利影响,造成细胞的机械损伤或抑制藻细胞的分裂增殖,不利于浮游植物群落形成[61]。青海湖水动力特征在平面上表现为:湖泊浅滨带为强水动力区域,越往湖心水动力越弱,甚至出现静水区域[62-63],相对稳定的水体环境促进了营养物质的运输和更新,有利于浮游植物生长繁殖,故青海湖显示出深水区的浮游植物丰度高于其他区域的趋势。从时间尺度上来看,夏秋两季的浮游植物丰度、生物量和优势类群及种类较春季均发生了较大的变化,春季丰度和生物量显著低于夏、秋两季,蓝藻门在夏、秋两季超越硅藻门成为最具优势的类群。这一变化与历史时期至今浮游植物群落的演变趋势类似,主要原因在于不同季节之间较大的水温变化。浮游植物的生长主要受温度调控,温度升高能直接增强浮游植物光合作用和呼吸作用,从而提高生长率及繁殖速率,而蓝藻门对光强和温度的要求高于硅藻门[47]。这也是夏、秋季蓝藻门的环离鞘丝藻成为第一优势种的原因。
本研究显示青海湖浮游植物多样性指数普遍较低,反映了青海湖浮游植物群落结构较为简单,对外部环境改变和内部群落变动的抵抗能力较小,这可能是由青海湖高海拔半咸水湖泊的特点所决定的。根据重复测量方差分析结果,各多样性指数受季节影响较大。在时间尺度上,Shannon-Wiener多样性和Margalef多样性指数均呈现先下降后上升的趋势。总体上,春季浮游植物多样性指数较高,可能是由于春季气温低,人类活动等外源干扰对水体影响较小,浮游植物群落结构较为稳定。然而,生物多样性与群落稳定性的关系较为复杂[64],本研究的调查时间有限,具体变化趋势背后的原因有待于在更大时空尺度上的进一步研究。总体来看,本研究发现青海湖浮游植物群落空间变化远小于季节变化,这与环境因子在空间和季节上的差异性检验结果一致。浮游植物群落结构变化是水体中物理、化学和生物因素共同作用的结果[65]。本研究中,大多数环境因子在季节间的差异均远大于区域间差异,尤其是盐度、水温和总磷等影响浮游植物群落的关键环境因子,因此这可能是造成浮游植物季节间差异比区域间差异更为显著的主要原因。
3.3 青海湖浮游植物与水环境之间的关系浮游植物通常是水体生境改变最先和最直接的反映者,其生长特性和群落结构组成与环境因子有密切的相关性[66]。本研究显示,影响青海湖浮游植物群落结构的主要环境驱动因子为盐度、水温和总磷。
水温不仅是影响浮游植物生长繁殖中重要的环境因子,也是影响浮游植物群落结构组成变化的主要因素[67-68]。与历史时期到当前浮游植物的演替趋势相似,青海湖浮游植物从春季到夏、秋季也呈现出硅藻占优—蓝藻占优的变化趋势,季节间水温的变化是造成这一变化的主要原因之一。青海湖春季温度低,硅藻门占据主要优势。随着时间推移,水温也随之上升,蓝藻门逐渐代替硅藻门成为第一优势类群。
水体中的营养物质是浮游植物生长和繁殖的重要物质基础,其浓度的时空变化会显著影响浮游植物的生物量以及群落结构的时空演变[69]。营养盐浓度升高通常会导致浮游植物丰度升高和物种组成的变化。青海湖磷的缺乏限制了浮游植物的生长繁殖。而夏、秋季水体中磷浓度的上升,促使浮游植物中蓝藻门的优势度大幅度增加[70],尤其在低温环境中,磷浓度的上升对浮游植物的生长速率有非常明显的促进作用[71]。因此,本研究中夏、秋两季浮游植物总丰度高于春季且优势类群为蓝藻门。
dbRDA分析显示盐度是影响青海湖浮游植物群落结构的第一驱动因子(P= 0.001)。盐度可以通过改变水体的溶解氧、渗透压等影响浮游植物的生长[72-73]。研究显示,蓝藻受盐度变化的影响程度大于硅藻,且更偏好低盐环境[74-75]。在本研究中,由于降雨和入湖河流的补给,夏、秋季样点盐度均低于10 ‰。尽管相比春季,盐度下降程度并不是非常大(约1.5 ‰ ~2 ‰),但这也可能是导致蓝藻门在夏、秋季丰度增加的原因之一。然而,盐度的季节变化是否确实在青海湖浮游植物群落演替中起着至关重要的作用,还需要今后的研究进一步验证。
4 结论青海湖浮游植物群落结构在时空尺度上有显著差异:空间上,深水区丰度显著高于其他区域,进水区物种数、Shannon-Wiener多样性指数和Margalef指数也高于其他区域。浮游植物在季节间的差异则更为显著:从春季到秋季,浮游植物丰度、生物量和物种数均呈现上升趋势,最具优势类群及种类也从春季的硅藻转变为夏、秋季的蓝藻;春季浮游植物Pielou均匀度和Simpson指数显著高于夏、秋季,秋季Margalef指数和Shannon-Wiener多样性指数高于夏季。与历史时期相比,本研究中浮游植物丰度显著增加,浮游植物群落结构呈现硅藻门—蓝藻门转变。盐度、水温和总磷是影响青海湖浮游植物生长的主要环境驱动因子。
青海湖作为我国重要的生态保护区,总体来说,目前其水生态环境健康状况仍处于较好水平。但在后续的管理保护中,需要进一步关注气候变化和人类活动对水生态环境带来的影响。未来应该警惕由于蓝藻过度繁殖带来的水华风险,开展关于草场淹没带来的营养盐释放和流域污染负荷输入等问题的研究,制定有效的管控措施,保障青海湖生态健康。
5 附录附表Ⅰ~Ⅴ见电子版(10.18307/2023.0212)。
| 附表Ⅰ 青海湖3个季节4个区域物理化学特征及Kruskal-Wallis分析结果* Appendix Ⅰ Physical and chemical characteristics of the 4 regions in 3 seasons and Kruskal–Wallis results in the Lake Qinghai (Those P-values < 0.05 are bold) |
| 附表Ⅱ 青海湖浮游植物属相对丰度的均值±标准差* Appendix Ⅱ Mean values ± SD of relative abundance of genera of phytoplankton in the Lake Qinghai |
| 附表Ⅲ 青海湖4个区域3个季节间浮游植物物种数、丰度、生物量和多样性指数的重复测量方差分析结果* Appendix Ⅲ Repeated measures ANOVA of effects of 4 regions among 3 seasons in on taxa richness, abundance, biomass, and diversity indices in the Lake Qinghai |
| 附表Ⅳ 区域两两对比* Appendix Ⅳ Pair-wise tests among regions |
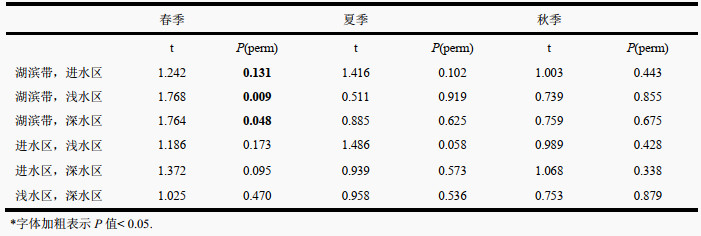
| 附表Ⅴ 季节两两对比* Appendix Ⅴ Pair-wise tests among seasons |
| [1] |
Meng XL, Jiang XM, Xiong X et al. Mediated spatio-temporal patterns of macroinvertebrate assemblage associated with key environmental factors in the Qinghai Lake area, China. Limnologica, 2016, 56: 14-22. DOI:10.1016/j.limno.2015.11.005 |
| [2] |
Zhu LP, Ju JT, Qiao BJ et al. Recent Lake changes of the Asia Water Tower and their climate response: Progress, problems and prospects. Chinese Science Bulletin, 2019, 64(27): 2796-2806. [朱立平, 鞠建廷, 乔宝晋等. "亚洲水塔"的近期湖泊变化及气候响应: 进展、问题与展望. 科学通报, 2019, 64(27): 2796-2806. DOI:10.1360/TB-2019-0185] |
| [3] |
Tolotti M, Manca M, Angeli N et al. Phytoplankton and zooplankton associations in a set of alpine high altitude lakes: Geographic distribution and ecology. Hydrobiologia, 2006, 562(1): 99-122. DOI:10.1007/s10750-005-1807-8 |
| [4] |
Lin QQ, Xu L, Hou JZ et al. Responses of trophic structure and zooplankton community to salinity and temperature in Tibetan lakes: Implication for the effect of climate warming. Water Research, 2017, 124: 618-629. DOI:10.1016/j.watres.2017.07.078 |
| [5] |
Wang TJ, Zhao LJ, Zhang X. Discussion on the method of ecotope comprehensive evaluation for Tibet plateau ecological shelter zone. Bulletin of Surveying and Mapping, 2018(9): 112-116. [王铁军, 赵礼剑, 张溪. 青藏高原生态屏障区生态环境综合评价方法探讨. 测绘通报, 2018(9): 112-116.] |
| [6] |
Yao TD, Chen FH, Cui P et al. From Tibetan Plateau to third pole and pan-third pole. Bulletin of Chinese Academy of Sciences, 2017, 32(9): 924-931. [姚檀栋, 陈发虎, 崔鹏等. 从青藏高原到第三极和泛第三极. 中国科学院院刊, 2017, 32(9): 924-931. DOI:10.16418/j.issn.1000-3045.2017.09.001] |
| [7] |
Diego F, Yamila B, Gisela M et al. Controlling factors in planktonic communities over a salinity gradient in high-altitude lakes. Annales De Limnologie-International Journal of Limnology, 2015, 51(3): 261-272. DOI:10.1051/limn/2015020 |
| [8] |
Williamson CE, Dodds W, Kratz TK et al. Lakes and streams as sentinels of environmental change in terrestrial and atmospheric processes. Frontiers in Ecology and the Environment, 2008, 6(5): 247-254. DOI:10.1890/070140 |
| [9] |
Catalan J, Camarero L, Felip M et al. High Mountain lakes: Extreme habitats and witnesses of environmental changes. Limnetica, 2006, 25(1): 551-584. DOI:10.23818/limn.25.38 |
| [10] |
Cartuche A, Guan ZY, Ibelings B et al. Phytoplankton diversity relates negatively with productivity in tropical high-altitude lakes from southern Ecuador. Sustainability, 2019, 11: 5235. DOI:10.3390/su11195235 |
| [11] |
Alcocer J, Lugo A, Fernández R et al. 20 years of global change on the limnology and plankton of a tropical, high-altitude lake. Diversity, 2022, 14(3): 190. DOI:10.3390/d14030190 |
| [12] |
Wang H, Yang SP, Fang SZ et al. Canonical correspondence analysis of relationship between characteristics of phytoplankton community and environmental factors in Dianchi Lake. China Environmental Science, 2016, 36(2): 544-552. [王华, 杨树平, 房晟忠等. 滇池浮游植物群落特征及与环境因子的典范对应分析. 中国环境科学, 2016, 36(2): 544-552. DOI:10.3969/j.issn.1000-6923.2016.02.034] |
| [13] |
Reynolds CS. The ecology of phytoplankton. Cambridge: Cambridge University Press, 2006. DOI:10.1017/cbo9780511542145
|
| [14] |
Wang YB, Hou ZY, Ye BB et al. The characteristics of spatial and temporal variations of phytoplankton in Poyang Lake and their influencing factors. Acta Scientiae Circumstantiae, 2015, 35(5): 1310-1317. [王艺兵, 侯泽英, 叶碧碧等. 鄱阳湖浮游植物时空变化特征及影响因素分析. 环境科学学报, 2015, 35(5): 1310-1317. DOI:10.13671/j.hjkxxb.2014.1052] |
| [15] |
Yan GH, Yin XY, Wang X et al. Effects of environmental factors on the composition of phytoplankton community in Sankou of the Yangtze River and the Western Dongting Lake. China Environmental Science, 2019, 39(6): 2532-2540. [严广寒, 殷雪妍, 汪星等. 长江三口-西洞庭湖环境因子对浮游植物群落组成的影响. 中国环境科学, 2019, 39(6): 2532-2540. DOI:10.3969/j.issn.1000-6923.2019.06.036] |
| [16] |
Liu XL, Hou WG, Dong HL et al. Distribution and diversity of cyanobacteria and eukaryotic algae in Qinghai-Tibetan lakes. Geomicrobiology Journal, 2016, 33(10): 860-869. DOI:10.1080/01490451.2015.1120368 |
| [17] |
Hillman AL, Abbott MB, Yu JQ. Climate and anthropogenic controls on the carbon cycle of Xingyun Lake, China. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 501: 70-81. DOI:10.1016/j.palaeo.2018.04.012 |
| [18] |
Sun X, Zhu GW, Yang WB et al. Spatio-temporal variations in phytoplankton community in Shahe reservoir, Tianmuhu, China. Environmental Science, 2017, 38(10): 4160-4168. [孙祥, 朱广伟, 杨文斌等. 天目湖沙河水库浮游植物群落结构的时空异质性. 环境科学, 2017, 38(10): 4160-4168. DOI:10.13227/j.hjkx.201704161] |
| [19] |
Wang ZF, Zhang W, Yang L et al. Characteristics of phytoplankton community and its relationship with environmental factors in different regions of Yilong Lake, Yunnan Province, China. Environmental Science, 2019, 40(5): 2249-2257. [王振方, 张玮, 杨丽等. 异龙湖不同湖区浮游植物群落特征及其与环境因子的关系. 环境科学, 2019, 40(5): 2249-2257. DOI:10.13227/j.hjkx.201810157] |
| [20] |
Meng XL, He YB, Song ZY et al. Struture and spatial distribution patterns of macrozoobenthos in Qinghai Lake area. Acta Hydrobiologica Sinica, 2014, 38(5): 819-827. [孟星亮, 何玉邦, 宋卓彦等. 青海湖区大型底栖动物群落结构与空间分布格局. 水生生物学报, 2014, 38(5): 819-827. DOI:10.7541/2014.123] |
| [21] |
Hao MY, Zhu H, Xiong X et al. Analysis on the distribution and origin of Cladophora in the nearshore water of Qinghai Lake. Acta Hydrobiologica Sinica, 2020, 44(5): 1152-1158. [郝美玉, 朱欢, 熊雄等. 青海湖刚毛藻分布特征变化及成因分析. 水生生物学报, 2020, 44(5): 1152-1158. DOI:10.7541/2020.133] |
| [22] |
Tang LY, Duan XF, Kong FJ et al. Influences of climate change on area variation of Qinghai Lake on Qinghai-Tibetan Plateau since 1980s. Scientific Reports, 2018, 8: 7331. DOI:10.1038/s41598-018-25683-3 |
| [23] |
Shen J, Liu X, Ryo M et al. A high-resolution climatic change since the Late Glacial Age inferred from multi-proxy of sediments in Lake Qinghai. Science in China Series D: Earth Sciences, 2005, 48(6): 742-751. DOI:10.1360/03yd0148 |
| [24] |
Sun YS, Li QJ, Liu T et al. Analysis of Qinghai Lake's water level changes and water balance analysis from 1956 to 2019. Journal of China Hydrology, 2021, 41(5): 91-96. [孙永寿, 李其江, 刘弢等. 青海湖1956—2019年水位变化原因及水量平衡分析研究. 水文, 2021, 41(5): 91-96.] |
| [25] |
Jin ZD, Zhang F, Wang HL et al. The reasons of rising water level in Lake Qinghai since 2005. Journal of Earth Environment, 2013, 4(3): 1355-1362. [金章东, 张飞, 王红丽等. 2005年以来青海湖水位持续回升的原因分析. 地球环境学报, 2013, 4(3): 1355-1362. DOI:10.7515/JEE201303008] |
| [26] |
Cheng J, Liu CH, Liu K et al. Potential impact of the dramatical expansion of Lake Qinghai on the habitat facilities and grassland since 2004. J Lake Sci, 2021, 33(3): 922-934. [程俭, 刘昌华, 刘凯等. 2004年以来青海湖快速扩张对人居设施与草地的潜在影响. 湖泊科学, 2021, 33(3): 922-934. DOI:10.18307/2021.0325] |
| [27] |
Gao LM, Zhang LL. Spatiotemporal dynamics of the vegetation coverage in Qinghai Lake Basin. Journal of Geo-Information Science, 2019, 21(9): 1318-1329. [高黎明, 张乐乐. 青海湖流域植被盖度时空变化研究. 地球信息科学学报, 2019, 21(9): 1318-1329. DOI:10.12082/dqxxkx.2019.180696] |
| [28] |
Chen FJ, Fu SY, Ma M et al. Effects of hypoxia stress on the activity of antioxidant enzymes, neuronal apoptosis and expression of related genes of telencephalon in Gymnocypris przewalskii. Acta Hydrobiologica Sinica, 1-12. [陈付菊, 付生云, 马敏等. 低氧胁迫对青海湖裸鲤端脑抗氧化酶活性、细胞凋亡及相关基因表达的影响. 水生生物学报, 1-12.] |
| [29] |
Tan XC, Shi JQ, Zhang H et al. Hydroacoustic assessment of fish resources in the Lake Qinghai with EY60 echosounder. J Lake Sci, 2009, 21(6): 865-872. [谭细畅, 史建全, 张宏等. EY60回声探测仪在青海湖鱼类资源量评估中的应用. 湖泊科学, 2009, 21(6): 865-872. DOI:10.18307/2009.0617] |
| [30] |
青海省生物研究所. 青海湖地区的鱼类区系和青海湖裸鲤的生物学. 北京: 科学出版社, 1975.
|
| [31] |
中国科学院兰州地质研究所等. 青海湖综合考察报告. 北京: 科学出版社, 1994.
|
| [32] |
Yang JX, Qi HF, Shi JQ et al. Survey of aquatic organisms in Lake Qinghai in summer. Qinghai Science and Technology, 2008, 15(6): 19-25. [杨建新, 祁洪芳, 史建全等. 青海湖夏季水生生物调查. 青海科技, 2008, 15(6): 19-25. DOI:10.3969/j.issn.1005-9393.2008.06.008] |
| [33] |
Yao WZ, Shi JQ, Qi HF et al. Study on the phytoplankton in Qinghai Lake during summer of 2006-2010. Freshwater Fisheries, 2011, 41(3): 22-28. [姚维志, 史建全, 祁洪芳等. 2006—2010年夏季青海湖浮游植物研究. 淡水渔业, 2011, 41(3): 22-28.] |
| [34] |
姚维志. 青海湖夏季饵料生物资源研究[学位论文]. 重庆: 西南大学, 2011.
|
| [35] |
国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法(第四版). 北京: 中国环境科学出版社, 2002.
|
| [36] |
胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态. 北京: 科学出版社, 2006.
|
| [37] |
韩茂森, 束蕴芳. 中国淡水生物图谱. 北京: 海洋出版社, 1995.
|
| [38] |
Burks AW, Shannon CE, Weaver W. The mathematical theory of communication. The Philosophical Review, 1951, 60(3): 398. DOI:10.2307/2181879 |
| [39] |
Simpson EH. Measurement of diversity. Nature, 1949, 163(4148): 688. DOI:10.1038/163688a0 |
| [40] |
Margalef R. Information theory in biology. General Systems Yearbook, 1958, 3: 36-71. |
| [41] |
Pielou EC. Species-diversity and pattern-diversity in the study of ecological succession. Journal of Theoretical Biology, 1966, 10(2): 370-383. DOI:10.1016/0022-5193(66)90133-0 |
| [42] |
Pan CM, Liu Y, An RZ et al. Phytoplankton in the Mitika Wetland, Tibet, China: 1. Spatio-temporal niche of dominant species. J Lake Sci, 2021, 33(6): 1805-1819. [潘成梅, 刘洋, 安瑞志等. 西藏麦地卡湿地的浮游植物——1.优势种的时空生态位. 湖泊科学, 2021, 33(6): 1805-1819. DOI:10.18307/2021.0616] |
| [43] |
Borcard D, Gillet F, Legendre P eds. Numerical ecology with R. New York: Springer, 2011. DOI:10.1007/978-1-4419-7976-6
|
| [44] |
Jiang XM, Pan BZ, Song ZY et al. Do functional traits of chironomid assemblages respond more readily to eutrophication than taxonomic composition in Chinese floodplain lakes?. Ecological Indicators, 2019, 103: 355-362. DOI:10.1016/j.ecolind.2019.04.029 |
| [45] |
王一郎. 藏北湖泊和青海湖流域的蓝藻多样性研究[学位论文]. 武汉: 中国科学院水生生物研究所, 2020.
|
| [46] |
Li QJ. Variation characteristic of surface water temperature and its response to climate change in Qinghai Lake. Yellow River, 2018, 40(11): 25-29. [李其江. 青海湖表层水温变化特征及对气候变化的响应. 人民黄河, 2018, 40(11): 25-29.] |
| [47] |
Li CC, Feng WY, Chen HY et al. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu-a large freshwater lake in China. Environmental Pollution, 2019, 245: 1050-1057. DOI:10.1016/j.envpol.2018.11.007 |
| [48] |
Diao YM. Situation of water environment of Qinghai Lake and protection measures. Yangtze River, 2014, 45(18): 33-36. [刁玉美. 青海湖流域水环境状况分析及保护对策. 人民长江, 2014, 45(18): 33-36.] |
| [49] |
Wang TC, Lu LH, Liu GX et al. Analysis of lakeside wetland evolution and driving factors around Qinghai Lake. Journal of China Institute of Water Resources and Hydropower Research, 2020, 18(4): 274-283. [王天慈, 卢丽华, 刘国祥等. 青海湖湖滨湿地演变与驱动因素分析. 中国水利水电科学研究院学报, 2020, 18(4): 274-283.] |
| [50] |
Liu ZQ, Pan BZ, Han X et al. Water environmental characteristics and water quality assessment of lakes in Tibetan Plateau. Environmental Science, 2022, 1-16. [刘智琦, 潘保柱, 韩谞等. 青藏高原湖泊水环境特征及水质评价. 环境科学, 2022, 1-16. DOI:10.13227/j.hjkx.202111079] |
| [51] |
Zhang W, Sun J, Nie HT et al. Seasonal and spatial variations of nutrient and the response of phytoplankton in PRE and adjacent sea areas. Acta Ecologica Sinica, 2015, 35(12): 4034-4044. [张伟, 孙健, 聂红涛等. 珠江口及毗邻海域营养盐对浮游植物生长的影响. 生态学报, 2015, 35(12): 4034-4044.] |
| [52] |
Brookes JD, Carey CC. Resilience to blooms. Science, 2011, 334(6052): 46-47. DOI:10.1126/science.1207349 |
| [53] |
Rigosi A, Carey CC, Ibelings BW et al. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnology and Oceanography, 2014, 59(1): 99-114. DOI:10.4319/lo.2014.59.1.0099 |
| [54] |
Ma JR, Deng JM, Qin BQ et al. Progress and prospects on cyanobacteria bloom-forming mechanism in lakes. Acta Ecologica Sinica, 2013, 33(10): 3020-3030. [马健荣, 邓建明, 秦伯强等. 湖泊蓝藻水华发生机理研究进展. 生态学报, 2013, 33(10): 3020-3030.] |
| [55] |
Liu YP, Huang YC, Wang HL. Discussion on the influence of environmental factors on diatom growth. Science & Technology Information, 2009(33): 725, 648. [刘杨平, 黄迎春, 王鹤立. 浅谈环境因子对硅藻生长的影响. 科技信息, 2009(33): 725, 648.] |
| [56] |
Liu XZ, Li ZJ, Cao YC et al. Common species composition, quantity variation and dominant species of planktonic microalgae in low salinity culture ponds. South China Fisheries Science, 2009, 5(1): 9-16. [刘孝竹, 李卓佳, 曹煜成等. 低盐度养殖池塘常见浮游微藻的种类组成、数量及优势种群变动. 南方水产, 2009, 5(1): 9-16.] |
| [57] |
Cai J, Wang XY, Zeng BB et al. Effects of salinity, light intensity and temperature on the growth, ARA and EPA contents in diatom Cyclotella sp. SHOU-B108. Journal of Shanghai Ocean University, 2016, 25(3): 406-414. [蔡敬, 王星宇, 曾蓓蓓等. 盐度-光照强度-温度对小环藻Cyclotella sp. SHOU-B108生长及ARA和EPA含量的影响. 上海海洋大学学报, 2016, 25(3): 406-414.] |
| [58] |
Håkanson L, Bryhn AC, Hytteborn JK. On the issue of limiting nutrient and predictions of cyanobacteria in aquatic systems. Science of the Total Environment, 2007, 379(1): 89-108. DOI:10.1016/j.scitotenv.2007.03.009 |
| [59] |
Reynolds C. The ecology of freshwater phytoplankton. Cambridge: Cambridge University Press, 1984.
|
| [60] |
Han YH, Hao LW, Wang Q et al. Prevailing winds controlled hydrodynamic characteristics of Qinghai Lake. Yellow River, 2016, 38(3): 51-55. [韩元红, 郝乐伟, 王琪等. 盛行风控制下的青海湖水动力特征. 人民黄河, 2016, 38(3): 51-55.] |
| [61] |
Zhou J, Qin BQ. review of the effects of wind waves on plankton in lakes. Advances in Water Science, 2018, 29(2): 293-300. [周健, 秦伯强. 风浪对湖泊浮游生物影响的研究进展. 水科学进展, 2018, 29(2): 293-300.] |
| [62] |
李树同, 王琪, 郝乐伟等. 青海湖水动力特征及其与沉积充填响应. 见: 第十五届全国古地理学及沉积学学术会议. 中国四川成都, 2018: 584.
|
| [63] |
Zhang YF, Hu CL, Wang M et al. A quantitative sedimentary model for the modern lacustrine beach bar (Qinghai Lake, Northwest China). Journal of Paleolimnology, 2018, 59(2): 279-296. DOI:10.1007/s10933-016-9930-2 |
| [64] |
Tian W, Zhang HY, Wang ZY et al. Phytoplankton diversity effects on community biomass and temporal stability in Lake Nansihu. China Environmental Science, 2017, 37(1): 319-327. [田旺, 张化永, 王中玉等. 南四湖浮游植物多样性与群落生物量、时间序列稳定性关系. 中国环境科学, 2017, 37(1): 319-327.] |
| [65] |
Calijuri MC, dos Santos ACA, Jati S. Temporal changes in the phytoplankton community structure in a tropical and eutrophic reservoir (Barra Bonita, S.P.-Brazil). Journal of Plankton Research, 2002, 24(7): 617-634. DOI:10.1093/plankt/24.7.617 |
| [66] |
Lu JS, Yang Z, Zhang Y. Algae functional group characteristics in reservoirs and lakes with differenttrophic levels in northwestern semi-humid and semi-arid regions in China. Journal of Environmental Sciences, 2018, 64: 166-173. DOI:10.1016/j.jes.2017.06.011 |
| [67] |
Da WY, Zhu GW, Wu ZX et al. Long-term variation of phytoplankton community and driving factors in Qiandaohu Reservoir, southeast China. J Lake Sci, 2019, 31(5): 1320-1333. [笪文怡, 朱广伟, 吴志旭等. 2002—2017年千岛湖浮游植物群落结构变化及其影响因素. 湖泊科学, 2019, 31(5): 1320-1333. DOI:10.18307/2019.0522] |
| [68] |
Zhang CM, Zhu YX, Song GF et al. Spatiotemporal pattern of phytoplankton community structure and its determining factors in the channel of the middle route of South-to-North Water Diversion Project. J Lake Sci, 2021, 33(3): 675-686. [张春梅, 朱宇轩, 宋高飞等. 南水北调中线干渠浮游植物群落时空格局及其决定因子. 湖泊科学, 2021, 33(3): 675-686. DOI:10.18307/2021.0305] |
| [69] |
Szymczak-żyƚa M, Krajewska M, Winogradow A et al. Tracking trends in eutrophication based on pigments in recent coastal sediments. Oceanologia, 2017, 59(1): 1-17. DOI:10.1016/j.oceano.2016.08.003 |
| [70] |
O'Neil JM, Davis TW, Burford MA et al. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae, 2012, 14: 313-334. DOI:10.1016/j.hal.2011.10.027 |
| [71] |
Xu H, Paerl HW, Qin BQ et al. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnology and Oceanography, 2010, 55(1): 420-432. DOI:10.4319/lo.2010.55.1.0420 |
| [72] |
Bergesch M, Garcia M, Odebrecht C. Diversity and morphology of Skeletonema species in southern Brazil, southwestern Atlantic Ocean. Journal of Phycology, 2009, 45(6): 1348-1352. DOI:10.1111/j.1529-8817.2009.00743.x |
| [73] |
Dong GC, Sun LF, Li XQ et al. Community characteristics of phytoplankton in seawater multilevel comprehensive utilization ponds of Yellow River Delta. Fishery Modernization, 2019, 46(5): 46-54. [董贯仓, 孙鲁峰, 李秀启等. 黄河三角洲梯度利用海水浮游植物群落特征分析. 渔业现代化, 2019, 46(5): 46-54.] |
| [74] |
Su ZJ, Wang YJ, Dong ZJ et al. Response of phytoplankton assemblages to the water-sediment regulation in the adjacent sea of the Yellow River mouth. Acta Oceanologica Sinica, 2015, 37(4): 62-75. [苏芝娟, 王玉珏, 董志军等. 调水调沙后黄河口邻近海域浮游植物群落响应特征. 海洋学报, 2015, 37(4): 62-75.] |
| [75] |
Ren H, Tian T, Yang YF et al. Spatial and temporal distribution of phytoplankton community and its relationship with environment factors in Nansha's Rivers, Pearl River Estuary. Acta Ecologica Sinica, 2017, 37(22): 7729-7740. [任辉, 田恬, 杨宇峰等. 珠江口南沙河涌浮游植物群落结构时空变化及其与环境因子的关系. 生态学报, 2017, 37(22): 7729-7740.] |