 2023, Vol. 35
2023, Vol. 35 


(2: 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008)
(3: 皖江流域退化生态系统的恢复与重建省部共建协同创新中心, 芜湖 241002)
(2: Nanjing Institute of Geography and Limnology, State Key Laboratory of Lake Science and Environment, Chinese Academy of Sciences, Nanjing 210008, P. R. China)
(3: Collaborative Innovation Center of Recovery and Reconstruction of Degraded Ecosystem in Wanjiang Basin Co-founded by Anhui Province and Ministry of Education, Wuhu 241002, P. R. China)
湖滨带通常是指湖滨水陆交错带,为湖泊流域陆生生态系统与水生生态系统之间的过渡带,对维持湖泊水质优良和生态系统完整性具有重要作用[1]。相比于敞水区,湖滨带具有明显的水文和地形梯度、生境异质、物种丰富和生产力高等特点[2]。作为陆地和湖泊水体之间的过渡带,湖滨带具有重要的生态和服务功能,在消解与滞留污染物、稳固湖岸与维持清水稳态、支撑区域生物群落、生产生物质和保持物种多样性等方面发挥重要作用[3-5]。由于湖滨带各岸段的基底、地形、气候、水文和水动力条件等环境因子和岸上土地利用类型及人类活动存在交互影响,从而导致湖滨带的生境空间结构在横向、纵向和轴向上的圈层结构特点,因此,不同岸段生态系统的群落结构、密度和生物量的时空分布格局、生物多样性等会表现出一定的差异性[1]。
浮游动物是水生生态系统的重要组成部分,在渔业养殖、环境监测和生态评估中发挥着重要作用;它们分布广泛,虽然体型较小,但繁殖速率快,因此能够迅速占领环境中的空余生态位,并将初级生产力转化成次级消费者可利用的形式,故而在水生态系统的物质循环和能量传递过程中扮演着重要角色[6]。作为水生食物链的重要一环,浮游动物一方面对浮游植物、原生动物和细菌的丰度起到调控作用;另一方面,作为一些鱼虾幼苗优质的饵料来源[7],其群落结构在鱼类密度较高的水体中受到“下行效应”的显著影响。因此,无论是作为功能类群还是指示类群[8],凭借其对环境变化敏感和快速响应,浮游动物是湖泊生态学研究的重要对象。
洪泽湖为亚热带大型浅水调蓄型湖泊,位于淮河中下游,具有防洪、灌溉、调水、渔业、水运和水电等多种功能[9]。受水利工程调蓄影响,多年来洪泽湖水位变化呈现出夏季低冬季高的特征,这与江淮流域多数湖泊相异。洪泽湖也是典型的过水性湖泊,其水位变幅较大(> 2 m),入湖河流泥沙多且水质较差。洪泽湖湖滨带生境具有其特殊性和复杂性,地势低缓,地形复杂多样,湿地和平原、岗地镶嵌分布,水陆交错,湖岸曲折,湖汊众多[10]。随着洪泽湖周边经济的快速发展,湖滨带受人类活动影响加剧,农业面源污染负荷高;围垦严重,围网、圈圩面积分别高达324和183 km2,占湖泊面积的30.8%;泥沙淤积等导致湖滨带生态系统退化严重;出现水生植物分布面积锐减、水体富营养化甚至部分湖区存在蓝藻水华等问题[11-14]。目前大部分研究主要集中于洪泽湖敞水区不同空间区域和季节间的结构差异,以鱼类和底栖动物为研究对象较多,针对其湖滨带浮游动物的群落结构特征及其响应机制尚鲜有研究。为探究洪泽湖湖滨带浮游动物群落结构的空间格局,本研究分析了洪泽湖湖滨带浮游动物群落结构及驱动因素,使用冗余分析(redundancy analysis, RDA)探究了洪泽湖水文、水化学特征、物理生境和饵料浮游植物等环境因子对浮游动物的影响,以期为洪泽湖生态环境保护和水生资源管理提供科学依据。
1 材料与方法 1.1 研究区概况及采样点设置洪泽湖属于大型过水型湖泊,湖泊平均水位为12.37 m,面积1576.9 km2。全湖水域由北部的成子湖湾、西部的溧河湖湾和西南部的淮河湾组成[15]。成子湖区多为中小入湖河流,区域水体流动性较差,湖滨带环境相对稳定;溧河洼区有湿地保护区,水深较浅,有大量围网圈圩存在;淮河湾设有大量圈圩,水流从西部淮河流入,水质主要受入湖河流淮河影响;东部湖滨带为人工修建的硬质大堤,受人类活动影响较大。综合考虑以上因素,将洪泽湖湖滨带按自然形成的湖湾划分为5个部分(图 1),于2020年8月进行采样,围绕洪泽湖周边湖滨带设置56个采样点,其中东部湖岸16个样点,淮河口12个样点,溧河洼10个样点,成子湖西11个样点,东北湖岸7个样点。

|
图 1 洪泽湖湖滨带采样点设置 Fig.1 Location of sampling sites in the littoral zone of Lake Hongze |
使用YSI-EXO多参数水质监测仪现场测定记录水深(WD)、透明度(SD)、水温(WT)、溶解氧(DO)、电导率(EC)、pH值、浊度(NTU)等指标。利用5 L采水器从表层0.5 m下、中层和底层0.5 m以上分别采样并混合后取1 L,密封立即送回实验室,按照《湖泊富营养化调查规范》测定总氮(TN)、总磷(TP)、溶解性总氮(TDN)、溶解性总磷(TDP)、磷酸盐(PO43--P)、硝酸盐(NO3--N)、高锰酸盐指数(CODMn)、溶解有机碳(DOC)、悬浮物(SS)及叶绿素a(Chl.a)浓度[16]。
1.2.2 浮游动植物浮游植物采样时,分层采集水样混合后取1 L,加入10~15 mL鲁哥氏液固定,静置48 h后浓缩定容至30 mL,用于原生动物、轮虫和浮游植物样品的种类鉴定与计数。枝角类和桡足类采样时,使用5 L有机玻璃采水器采集表层0.5 m下层、中层和底层0.5 m以上水样各10 L,经25号浮游生物网过滤浓缩,加入甲醛溶液至终浓度为4%固定,回到实验室后定容到30 mL。每个样品计数3次,取平均值后计算分析其密度。大型浮游动物生物量的计算中,枝角类的生物量根据体长-体重回归方程换算[17],桡足类根据外形的几何形状换算为生物量[18];原生动物和轮虫用体积法求得生物体积,比重取l,再根据体积换算为生物量[17],浮游动物生物量都用湿重来表示。浮游动物种类鉴定参照文献[19-22]。藻类鉴定参照文献[23];由于浮游植物的比重接近1,藻类的生物量可以直接通过浮游植物的体积换算得到[24]。
1.2.3 其他环境因子基于逐日气象数据分析获得洪泽湖近2年盛行风向,计算湖泊内每个监测点东、南、西、北4个方位离岸距离,选取2个盛行风向下监测点离岸距离加和作为湖泊扰动指数,进而获得扰动指数的空间分布,表征风浪扰动强度的空间格局[25]。水生植物盖度采用目测法估计,以百分比方式表征,参照Braun-Blanquet多盖度方法。
1.3 数据分析 1.3.1 统计检验在R语言car包中,利用leveneTest对环境因子进行方差齐性检验,P>0.05表现为符合方差齐性,用单因素方差检验,若出现显著性差异(P < 0.05),使用agricolae包中LSD法进行两两比较;若不符合方差齐性(P < 0.05),进行kruskal.test非参数检验,若出现显著差异则通过bonferron校正的Mann-Whitney配对比较评估差异。
1.3.2 营养状态分析应用综合营养状态指数(TLI(Σ))评价水质营养状态,计算公式为:
| $ {TLI}(\Sigma)=\sum\limits_{j=1}^m W_j \cdot {TLI}(j) $ | (1) |
式中,Wj为第j种参数的营养状态指数相关权重;TLI(j)为第j种参数的营养状态指数,Chl.a、TP、TN、SD和CODMn的权重分别为0.267、0.188、0.179、0.183和0.183。
1.3.3 浮游动物多样性分析对湖滨带的浮游动物群落数据进行ANOSIM分析,检验不同区域的群落组成之间是否存在显著差异。利用Margalef丰富度指数(D)、Shannon-Wiener多样性指数(H′)和均匀度指数(J)、优势度指数(Y)分析洪泽湖湖滨带浮游动物群落多样性,计算公式如下:
| $ D=(S-1) / \ln N $ | (2) |
| $ H^{\prime}=-\sum\limits_{i=1}^s\left(N_i / N\right) \ln \left(N_i / N\right) $ | (3) |
| $ Y=\left(N_i / N\right) \times f_i $ | (4) |
式中, S为物种总数;N为所有种类的总个体数;Ni为第i种的个体数;fi为某物种出现的频率;以Y>0.02定为优势种。
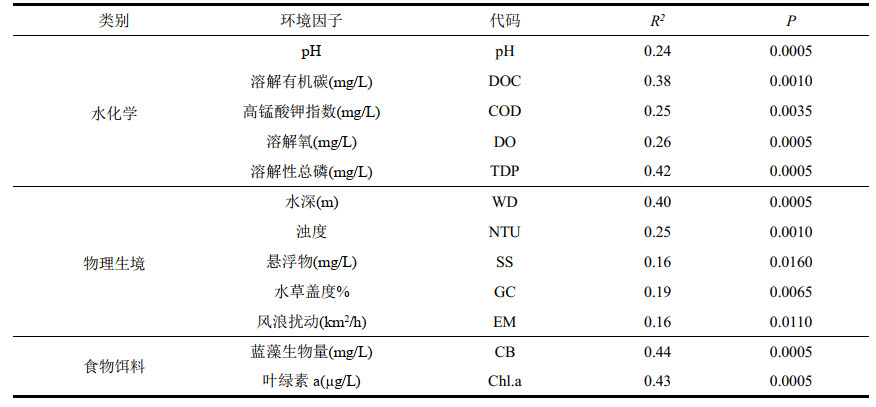
1.3.4 环境因子梯度分析在R语言中对洪泽湖湖滨带点位与环境因子进行主成分分析(principal component analysis, PCA);依据相对丰度大于1%,出现频率大于25%的标准对洪泽湖湖滨带的浮游动物进行筛选[26],筛选出18种主要种属的浮游动物。在去趋势对应分析(detrended correspondence analysis, DCA)中得到排序轴梯度长度最大值为2.823,所以选择冗余分析(redundancy analysis, RDA)。在分析过程中,将环境因子按性质分为水化学、物理生境和食物饵料三大部分[27]。水化学包括溶解氧、电导率、pH、TN、TP、DOC、COD、PO43--P、NO3--N;物理生境包括水深、透明度、浊度、悬浮物、水草盖度、风浪扰动;食物饵料有蓝藻、绿藻、甲藻、裸藻、硅藻和隐藻生物量。将经过前向选择(P < 0.05)筛选后的3组环境因子分别使用CANOCO for Windows 4.5对浮游动物物种数据与环境因子数据进行冗余分析。
1.3.5 影响因素分析方差分解是分析预测残差的标准差有不同信息的冲击影响的比例,亦即对应内生变量对标准差的贡献比例[28]。使用方差分解来反映水化学、物理生境和食物饵料对浮游动物密度分布及其群落多样性影响的相关程度,解析具体环境因子的解释程度。在vegan包中,使用ordiR2step进行环境因子筛选后利用varpart函数进行方差分解。
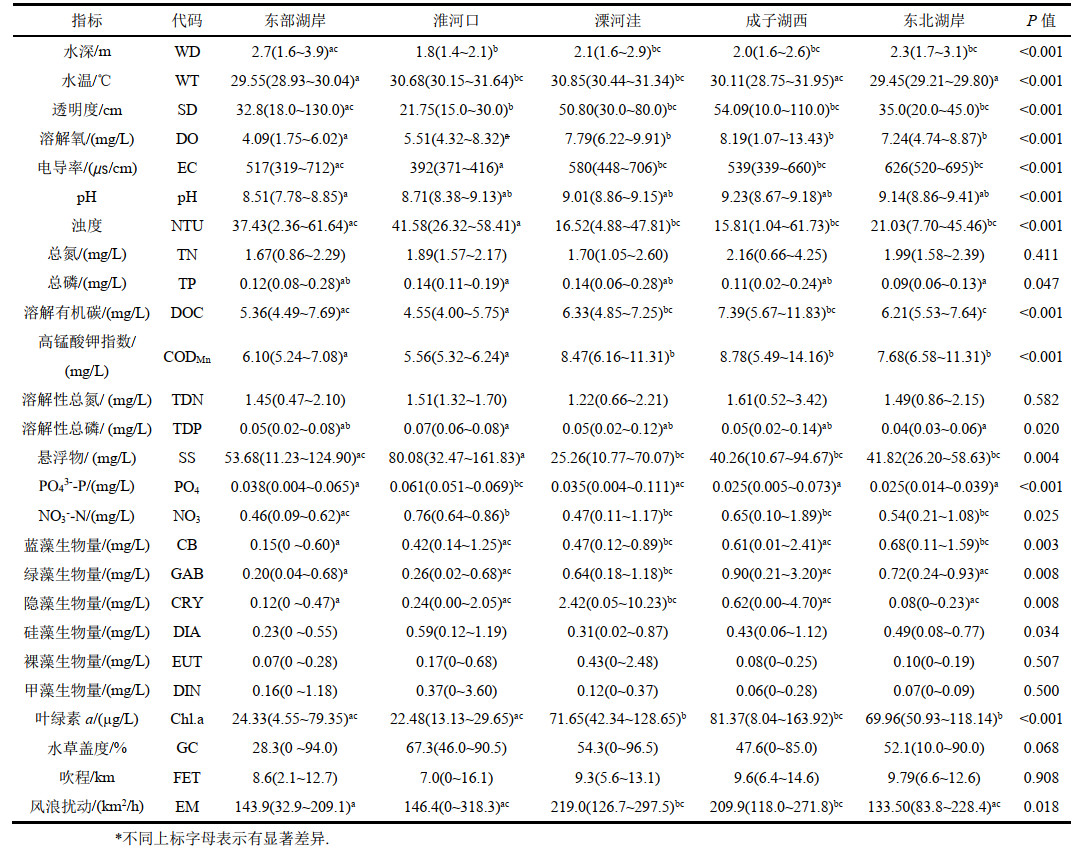
2 结果 2.1 洪泽湖湖滨带理化因子变化特征洪泽湖湖滨带8月份平均水深2.2 m,水温30.1℃,pH值为8.9,整体呈弱碱性,综合营养状态指数(TLI(Σ))均值为62.07,属于中度富营养状态。不同区域湖滨带TN、TDN浓度没有显著差异(附表Ⅰ);总磷浓度变化在0.02~0.28 mg/L之间,均值为0.12 mg/L,东部湖岸、溧河洼和成子湖西与淮河口和东北湖岸具有显著差异;pH东部湖岸显著低于其他湖区,均值为8.51;成子湖西和溧河洼浊度低,透明度较高,而淮河口悬浮物浓度最高,与其他湖区均具有显著差异,平均达到80.08 mg/L,透明度最低。DOC在溧河洼和成子湖西没有显著差异且高于其他湖区,分别达到6.33和7.39 mg/L;CODMn在东部湖岸和淮河口显著低于其他湖区,其中淮河口最低平均为5.56 mg/L。全湖滨带浮游植物中硅藻门、裸藻门和甲藻门生物量没有显著差异,成子湖西和东北湖岸蓝藻门和绿藻门生物量都较高,硅藻门生物量在成子湖西分布最高。洪泽湖湖滨带叶绿素a浓度平均为47.69 μg/L,最高的为成子湖西81.37 μg/L,最低为淮河口仅22.48 μg/L,不同湖区的藻类分布差异显著。风浪扰动表现为溧河洼与成子湖西显著高于其他湖区,而吹程和水草盖度无显著差异。
2.2 洪泽湖湖滨带的种类组成与优势种共鉴定出浮游动物52种,其中轮虫22种,占浮游动物总物种数的42.3%;原生动物15种,占28.8%;枝角类7种,占13.5%;桡足类8种,占15.4%。原生动物和轮虫是湖滨带浮游动物的主要种类(图 2)。

|
图 2 洪泽湖湖滨带浮游动物相对丰度 Fig.2 Zooplankton relative abundance in the littoral zone of Lake Hongze |
ANOSIM分析可得P=0.001,说明湖滨带不同区域之间的群落组成存在显著差异。以优势度Y>0.02为标准,优势种共有18种(表 1)。东部湖岸的拟铃虫属(Tintinnopsis sp.)、大弹跳虫(Halteria grandinella)和简弧象鼻溞(Bosmina coregoni)为其特有优势种;螺形龟甲轮虫(Keratella cochlearis)为淮河口的
特有优势种;东北湖岸特有优势种为镰状臂尾轮虫(Brachionus forficula)和尾突臂尾轮虫(B. caudatus)。溧河洼和成子湖西无特有优势种,溧河洼优势度较高的浮游动物种类有砂壳虫属(Diffllugia sp.)、萼花臂尾轮虫(B. calyciflorus)、曲腿龟甲轮虫(K. valga)和针簇多肢轮虫(Polyarthra trigla),成子湖西砂壳虫属(Diffllugia sp.)、剪形臂尾轮虫(B. forficula)和曲腿龟甲轮虫较高。浮游动物优势种在不同区域存在差异,其中砂壳虫属、曲腿龟甲轮虫在洪泽湖湖滨带各个区域都有分布且都占有优势,是洪泽湖湖滨带浮游动物的共有优势种。
| 表 1 洪泽湖湖滨带浮游动物优势种及优势度空间变化* Tab. 1 Spatial changes of dominant species and dominance degree of zooplankton in the littoral zone of Lake Hongze |
浮游动物的平均丰度为821.65 ind./L,原生动物和轮虫占比92.67%,枝角类和轮虫占比7.33%。平均生物量为1247.45 mg/L,原生动物占比0.18%,轮虫占比25.3%,枝角类占比69.92%,桡足类占比4.60%。成子湖西浮游动物丰度最高(1231.00 ind./L),溧河洼、东北湖岸、东部湖岸浮游动物丰度分别为892.35、745.50和731.97 ind./L,淮河口丰度最低(551.50 ind./L);浮游动物生物量分布最高的区域是东部湖岸(2511.07 mg/L)、成子湖西、东北湖岸、溧河洼浮游动物生物量分别为1146.61、668.46、605.45 mg/L,最低值在淮河,为527.62 mg/L(图 3)。

|
图 3 洪泽湖湖滨带浮游动物丰度及生物量 Fig.3 Zooplankton abundance and biomass in the littoral zone of Lake Hongze |
就具体的类群而言,湖滨带浮游动物丰度皆以原生动物和轮虫为主,占比85%以上;生物量的组成分布与丰度存在差异,东部湖岸枝角类的生物量最高(2276.37 mg/L),占90%以上;东北湖岸枝角类生物量占85%以上;淮河口浮游动物生物量以轮虫为主,占81.85%;溧河洼以轮虫(61.78%)和枝角类(24.72%)为主;成子湖西轮虫(42.50%)、枝角类(46.76%)和桡足类(12.79%)都有着较高的生物量(图 4、图 5)。从浮游动物丰度来看,成子湖西和溧河洼丰度高于其他湖区,原生动物和轮虫占据绝对优势;从生物量来看,枝角类和轮虫是构成湖滨带浮游动物生物量的主要类群,而原生动物和桡足类占比很低,其中枝角类在东部湖岸和东北湖岸处于绝对优势,轮虫是淮河口和溧河洼浮游动物的主要类群,成子湖西则以轮虫、枝角类和桡足类为主要生物量构成。

|
图 4 洪泽湖湖滨带浮游动物丰度(a)与生物量(b)空间变化 Fig.4 Spatial variation of zooplankton abundance (a) and biomass (b) in the littoral zone of Lake Hongze |

|
图 5 洪泽湖湖滨带浮游动物丰度(a)与生物量(b)及空间分布 Fig.5 Spatial distribution of zooplankton abundance (a) and biomass (b) in the littoral zone of Lake Hongze |
Shannon-Wiener多样性指数(H′)平均为1.64,指数最高值位于成子湖西(1.92),东部湖岸最低为1.34;Margalef丰富度指数(D)平均为1.39,溧河洼区域指数最高为1.68,东部湖岸最低为1.11;Pielou均匀度指数(J)平均为0.71,成子湖西指数最高为0.76,东部湖岸最低为0.64(图 6)。整体表现为,成子湖西和溧河洼H′、D和J相对较高,东北湖岸次之,淮河口和东湖湖岸最低。

|
图 6 洪泽湖湖滨带浮游动物群落多样性及空间变化 Fig.6 Spatial variation of zooplankton diversity indices in the littoral zone of Lake Hongze |
从排序轴上看(图 7),第一轴特征根值为6.24,第二轴特征根值为5.55,前两轴共解释了42.1%的环境因子差异。总氮、悬浮物、叶绿素a、COD、DOC、pH、蓝藻生物量和溶解氧是湖滨带主要的环境因子。分析表明,东部湖岸与电导率、DOC相关性较强;淮河口水体较深,浊度较高;溧河洼水质中存在大量颗粒悬浮物;成子湖西与东北湖岸受多种环境因子影响,且无大量悬浮物,浊度较低,具有多样的水环境。

|
图 7 洪泽湖湖滨带各样点与环境因子PCA排序(各环境因子代码见附表Ⅰ) Fig.7 PCA ordination of sampling sites and environmental factors in the littoral zone of Lake Hongze |
通过前向选择对环境因子进行筛选(P < 0.05),共筛选出12种对样点分布具有显著影响的环境因子(附表Ⅱ)。在RDA排序分析和方差分解分析中,将环境因子分为水化学、物理生境和食物饵料3部分。结果表明,在水化学排序轴中(图 8a),前两轴共解释了68.46%的水化学环境因子对浮游动物群落的影响程度。其中,针簇多肢轮虫和萼花臂尾轮虫与溶解氧浓度具有显著的正相关关系;圆筒异尾轮虫与pH、角突臂尾轮虫与DOC、螺形龟甲轮虫与水体中溶解性总磷、砂壳虫属与COD均呈高度正相关;水化学环境因子对于轮虫群落的影响较大。在物理生境排序轴中(图 8b),前两轴共解释了72.67%的物理生境环境因子对浮游动物群落的影响程度。大弹跳虫、拟铃虫属受水深的影响较高,风力扰动和水草盖度与大多数浮游动物呈正相关,悬浮物与大多数浮游动物呈负相关。在食物饵料排序轴中(图 8c),第一轴解释了80.78% 的食物饵料环境因子对浮游动物群落的影响程度,其中短棘裂足臂尾轮虫、镰状臂尾轮虫、萼花臂尾轮虫和螺形龟甲轮虫与蓝藻门生物量呈正相关。

|
图 8 洪泽湖湖滨带浮游动物主要种属与环境因子RDA排序 (a:水化学排序;b:物理生境排序;c:食物饵料排序)(SP1~SP18代码见表 1) Fig.8 RDA ordination of the primary zooplankton genus and environmental factors in the littoral zone of Lake Hongze (a: Water chemical ranking; b: Physical habitat ranking; c: Food ranking) |
方差分解结果表明,水化学因子DOC、PO43--P、COD对浮游动物丰度解释程度为17%,其与食物饵料的交互作用解释了14%(图 9);轮虫主要受到水化学环境因子及其与食物饵料的交互作用,总解释量为32%;枝角类受水深、浊度和悬浮物这些物理生境环境因子的影响,解释程度为22%,物理生境因子与水化学、食物饵料的交互作用总解释量为36%;磷酸盐和pH对桡足类丰度解释程度为6%,风浪扰动和悬浮物与水化学的交互作用贡献率为8%;枝角类和桡足类受食物饵料影响较小。Margalef指数主要受水化学环境因子及其与物理生境因子的交互作用影响;Shannon-Wiener指数主要受水体中悬浮物的影响及其与水化学、食物饵料的交互作用影响。

|
图 9 洪泽湖湖滨带环境因子方差分解结果 Fig.9 Result of variance partitioning of environmental factors in the littoral zone of Lake Hongze |
本次调查共发现洪泽湖湖滨带的浮游动物52种,空间上5个湖区浮游动物种类组成类似,群落结构均表现为以轮虫为主,甲壳动物次之,原生动物种类最少。其中东部湖岸浮游动物种类发现33种,溧河洼31种,成子湖西34种,东北湖岸26种,淮河口24种最少。都雪等[29]在2010年5月至2011年2月对洪泽湖浮游动物进行季度采样发现,主要优势种为螺形龟甲轮虫、前节晶囊轮虫、长肢多肢轮虫、萼花臂尾轮虫、曲腿龟甲轮虫、长额象鼻溞、透明溞、汤匙华哲水蚤、中华窄腹剑水蚤。陈业等[30]在2017年3月至2018年2月的逐月调查数据中发现,洪泽湖夏季优势种有矩形龟甲轮虫、曲腿龟甲轮虫、针簇多肢轮虫、简弧象鼻溞和角突网纹溞。本次调查与之相比,可以看出浮游动物群落的优势种因为水体环境等因素存在交叉和演替,随着生境的改变而产生不同的群落结构特征[31]。淮河口特有优势种螺形龟甲轮虫,东部湖岸特有优势种简弧象鼻溞,湖滨带广泛分布的优势种针簇多肢轮虫和臂尾轮属等皆为富营养水体的指示物种[32],表明洪泽湖水体处于富营养状态,且与水体理化指标分析结果相似,对水生态的健康状况具有一定的警示作用。
3.2 浮游动物分布特征及其受环境因子的影响国内已有的研究中,太湖滨岸带浮游动物分布主要受到溶解性总氮、总磷、透明度、溶解氧及pH值的影响[33];武汉东湖浮游动物分布受温度、溶解氧、pH、盐度和ORP的影响较大[34];鄱阳湖夏季影响浮游动物的环境因子为水深、电导率、浊度和溶解氧[35]。与其他浅水湖泊相比,本研究在RDA排序分析和方差分解分析中,轮虫丰度主要受到DOC、COD、PO43--P和叶绿素a的影响,枝角类丰度主要受到悬浮物、浊度、水深和硅藻的影响,桡足类丰度主要受到pH、PO43--P和风浪扰动、悬浮物的影响。
水体pH与浮游动物的分布有密切的联系。有学者发现,轮虫、桡足类更适宜生活在偏碱性水体中,枝角类则在酸性、中性及微碱性水体中生活的更好[36]。在本研究中东部湖岸的pH均值最低, 为8.51,成子湖西pH均值最高, 为9.23,这与东部湖岸高丰度枝角类,成子湖西高丰度轮虫和桡足类相呼应。成子湖西和溧河洼的Shannon-Wiener多样性指数超过1.75,均匀度指数超过1.50,淮河口和东部湖岸较低,进一步表明成子湖西和溧河洼的群落结构更为复杂和稳定[37]。
轮虫广泛分布于各类淡水水体中,其丰度通常可达1000 ind./L以上;食物充足条件下其丰度可超过5000 ind./L[38]。而本研究中的丰度都低于这一水平,原因其一可能是采样时间在8月份,处于洪泽湖的涨水期[39],尤其是湖体边缘的湖滨带受到了较为严重的影响,水位波动使得悬浮颗粒物增多,水体浊度增加。从排序图和方差分解中可看出,悬浮物是影响浮游动物物种分布和群落多样性的一个重要环境因子。有研究显示,曲腿龟甲轮虫的分布主要与水体浊度有关[33],这可能也是曲腿龟甲轮虫为洪泽湖湖滨带共有优势种的主要原因。方差分解中悬浮物对枝角类丰度解释程度较高,湖滨带枝角类生物量偏高,其受到的捕食压力较低[40]。其二是由于淮河口和溧河洼存在大量围网圈圩养殖,导致畜禽养殖的氮磷输入较高,养殖废弃物加剧湖泊水体富营养化,溧河洼浮游动物丰度较高。而淮河口流入携带氮、磷等污染物质的淮河水,水体具有大量的悬浮物,浊度较高[15]。淮河口和东部湖岸作为过水区,水流速度较快,而浮游动物游泳能力弱不足以抵抗水流,使得浮游动物丰度偏低;同时受到入湖河流(淮河)的影响,浊度偏高,其区域叶绿素a浓度低,影响了浮游动物的群落结构[41-43]。生物量方面,东部湖岸浮游动物生物量达到2511.07 μg/L,远高于东岸的淮河口527.62 μg/L和溧河洼605.45 μg/L。这与洪泽湖夏季盛行东南风,且西岸在增风效应作用下风速明显大于东岸,东部湖作为顺风站点,浮游动物尤其是枝角类生物量的顺风积累可能是主要原因之一[44-45]。
在RDA排序中,镰状臂尾轮虫、短棘裂足臂尾轮虫、萼花臂尾轮虫和螺形龟甲轮虫与叶绿素a浓度和蓝藻生物量呈正相关。薛庆举等[46]在洪泽湖蓝藻中普遍检测出较低浓度的藻毒素,萼花臂尾轮虫能抵抗藻毒素并从中获取营养[47];Liu等[48]围隔实验发现,螺形龟甲轮虫受微囊藻水华的影响较小,几乎都是随着蓝藻水华的出现而出现并大量繁殖。已有研究发现,在食物竞争中大型枝角类常是优胜者,而蓝藻能显著降低枝角类的个体大小,减弱枝角类对轮虫的竞争抑制[49],从而影响浮游动物的群落结构,间接提高浮游动物群落中轮虫的相对重要性[50]。螺形龟甲轮虫与TDP呈显著正相关,萼花臂尾轮虫与叶绿素a浓度呈正相关,体现其为富营养水体的指示种[32]。水生植物为浮游动物提供庇护所,大部分浮游动物与水草盖度呈显著正相关。通过对物种与环境的RDA排序可以更加直观地展现出物种的所适生境,以及影响不同物种分布的主要环境变量。
3.3 洪泽湖湖滨带浮游动物群落与湖心区比较洪泽湖湖滨带呈现向藻型浊水态转换的趋势,水体营养盐浓度高,浊度高,透明度低,水生植物稀疏,浮游藻类占据优势[51]。洪泽湖湖滨带较湖心区而言,取同时期敞水区10个样点作为对比(未发表数据),在水质方面,湖心区平均叶绿素a浓度(35.82 μg/L)、总氮浓度(1.39 mg/L)、总磷浓度(0.11 mg/L)、高锰酸盐指数(5.5 mg/L)、透明度(22.25 cm)等低于湖滨带,溶解氧浓度(8.13 mg/L)、水深(2.51 m)高于湖滨带。在浮游动物群落结构方面,湖心区浮游动物平均密度(1036.2 ind./L)和平均生物量(1939.7 μg/L)均高于湖滨带,与以往研究较为一致[29, 30, 52]。湖心区优势种为砂壳虫属、拟铃虫属、圆筒异尾轮虫(Trichocerca cylindrica)和简弧象鼻溞,比湖滨带5个湖区优势种结构简单;平均Shannon-Wiener指数(1.40)、Margalef丰富度指数(1.36)和Pielou均匀度指数(0.60)都略高于湖心区。
溧河洼、东北湖岸等都存在大量的围网养殖,受到人类活动的影响大于湖心区。该区域养殖污染物氮磷的输入满足了浮游植物的生长,藻类生物量积累为浮游动物密度增长提供了充足的食物饵料[53];而淮河口和东部湖岸悬浮物浓度高,水体浊度大,影响浮游植物光合作用速率,导致浮游植物生物量的降低[54],该区域浮游动物的丰度较低。湖滨带受到风浪扰动、富营养化等多种因素影响,空间异质性强,增强了浮游动物的代谢,不利于浮游动物的生长量累积[55];浮游动物趋于小型化和结构简单化,形成了洪泽湖湖滨带以小型浮游动物为主,丰富度指数和均匀度指数较低的结构特征。这种结构特征也与湖心区类似,但湖滨带要更为明显。从浮游动物结构特征层面来看,恢复洪泽湖湖滨带草型生态系统,对维持浮游动物群落多样性、丰富性和结构稳定性具有重要意义。
4 结语本次调查鉴定出浮游动物共52种,其中轮虫22种、原生动物15种、枝角类7种、桡足类8种。群落结构丰度表现为以原生动物和轮虫为主,生物量以轮虫和枝角类为主。在空间分布上,成子湖西的浮游动物丰度、生物量和群落多样性指数都较高,溧河洼次之,群落结构较为稳定;过水区淮河口浮游动物丰度、生物量和群落多样性指数较低,群落结构较不稳定。洪泽湖湖滨带浮游动物的群落结构主要受到颗粒悬浮物、叶绿素a和富营养化等因素的影响。本文对比发现湖滨带比敞水区受人类活动影响更大,水质较差,浮游动物群落多样性较低,结构更为简单,而国内对于湖滨带的研究较少,其作为湖岸缓冲带的关键作用应得到进一步重视。建议加强洪泽湖湖滨带的生态管理,如削减颗粒悬浮物含量和降低富营养化水平,恢复湖滨带草型生态系统,对维持浮游生物群落结构稳定具有重要意义。
5 附录附表Ⅰ和Ⅱ见电子版(DOI: 10.18307/2023.0218)。
| 附表Ⅰ 洪泽湖湖滨带不同区域环境因子特征 Appendix Ⅰ Characteristics of environmental factors in different regions in littoral zone of Lake Hongze |
| 附表Ⅱ 环境因子筛选结果 Appendix Ⅱ Results of the selection on environmental variables |
| [1] |
Ye C, Li CH, Deng TT. Structures and ecological functions of lake littoral zones. Research of Environmental Sciences, 2015, 28(2): 171-181. [叶春, 李春华, 邓婷婷. 论湖滨带的结构与生态功能. 环境科学研究, 2015, 28(2): 171-181. DOI:10.13198/j.issn.1001-6929.2015.02.02] |
| [2] |
Ostendorp W, Schmieder K, Jöhnk K. Assessment of human pressures and hydromorphological impacts on lakeshores in Europe. International Journal of Ecohydrology & Hydrobiology, 2004, 4(4): 379-395. |
| [3] |
Wantzen KM, Junk WJ, Rothhaupt KO. An extension of the flood pulse concept (FPC) for lakes. Hydrobiologia, 2008, 613: 151-170. DOI:10.1007/s10750-008-9480-3 |
| [4] |
Schmieder K. European lake shores in danger—concepts for a sustainable development. Limnologica, 2004, 34(1/2): 3-14. DOI:10.1016/S0075-9511(04)80016-1 |
| [5] |
Wang HZ. Basic concepts of lakeshore zones(preface). Resources and Environment in the Yangtze Basin, 2012, 21(S2): 1-2. [王洪铸. 湖滨带的基本概念(代前言). 长江流域资源与环境, 2012, 21(S2): 1-2.] |
| [6] |
Offem BO, Samsons YA, Omoniyi IT et al. Dynamics of the limnological features and diversity of zooplankton populations of the Cross River System SE Nigeria. Knowledge and Management of Aquatic Ecosystems, 2009, 393: 02. DOI:10.1051/kmae/2009013 |
| [7] |
Yang YF, Huang XF. Advances in ecological studies on zooplankton. J Lake Sci, 2000, 12(1): 81-90. [杨宇峰, 黄祥飞. 浮游动物生态学研究进展. 湖泊科学, 2000, 12(1): 81-90. DOI:10.18307/2000.0113] |
| [8] |
Duggan IC, Green JD, Shiel RJ. Distribution of rotifers in North Island, New Zealand, and their potential use as bioindicators of lake trophic state. Hydrobiologia, 2001, 446/447: 155-164. DOI:10.1007/978-94-010-0756-6_22 |
| [9] |
Yang GL, Han AM, Liu YK et al. The relationship of eutrophication of the lake of Hongze and environmental physical-chemical factors. The Administration and Technique of Environmental Monitoring, 2003, 15(2): 17-20. [杨广利, 韩爱民, 刘轶琨等. 洪泽湖富营养化与环境理化因子间的关系. 环境监测管理与技术, 2003, 15(2): 17-20. DOI:10.3969/j.issn.1006-2009.2003.02.008] |
| [10] |
Yang SJ. Discussion on the ecological construction of the western lakeshore of Hongze Lake. Chongqing Environmental Science, 2003, 25(11): 91-92, 102. [杨士建. 对洪泽湖西部湖滨生态建设的探讨. 重庆环境科学, 2003, 25(11): 91-92, 102.] |
| [11] |
Zhang CW, Zhang TL, Zhu TB et al. Community structure of macrozoobenthos and its relationship with environmental factors in lake Hongze. Journal of Hydroecology, 2012, 33(3): 27-33. [张超文, 张堂林, 朱挺兵等. 洪泽湖大型底栖动物群落结构及其与环境因子的关系. 水生态学杂志, 2012, 33(3): 27-33. DOI:10.15928/j.1674-3075.2012.03.003] |
| [12] |
Cai YJ, Zhang Z, Tang RG et al. Evaluation and protection of ecosystem health in Hongze Lake. Jiangsu Water Resources, 2020(7): 1-7, 13. [蔡永久, 张祯, 唐荣桂等. 洪泽湖生态系统健康状况评价和保护. 江苏水利, 2020(7): 1-7, 13.] |
| [13] |
Hu GF, Yang SJ. Biodiversity conservation and sustainable utilization in the western lakeside of Hongze Lake. Yunnan Environmental Science, 2004, 23(2): 34-38. [胡桂芳, 杨士建. 洪泽湖西部湖滨的生物多样性保护和可持续利用. 云南环境科学, 2004, 23(2): 34-38. DOI:10.3969/j.issn.1673-9655.2004.02.010] |
| [14] |
Yan CZ, Jin XC, Zhao JZ et al. Ecological restoration and reconstruction of degraded lakeside zone ecosystem. Chinese Journal of Applied Ecology, 2005, 16(2): 360-364. [颜昌宙, 金相灿, 赵景柱等. 湖滨带退化生态系统的恢复与重建. 应用生态学报, 2005, 16(2): 360-364. DOI:10.13287/j.1001-9332.2005.0448] |
| [15] |
Li Y, Zhang Z, Cheng JH et al. Water quality change and driving forces of Lake Hongze from 2012 to 2018. J Lake Sci, 2021, 33(3): 715-726. [李颖, 张祯, 程建华等. 2012——2018年洪泽湖水质时空变化与原因分析. 湖泊科学, 2021, 33(3): 715-726. DOI:10.18307/2021.0308] |
| [16] |
金相灿, 屠清瑛. 湖泊富营养化调查规范. 北京: 中国环境科学出版社, 1990.
|
| [17] |
赵文. 水生生物学. 北京: 中国农业出版社, 2005.
|
| [18] |
Chen XM. Biomass calculation of freshwater copepoda. Acta Hydrobiologica Sinica, 1981, 47(3): 397-408. [陈雪梅. 淡水桡足类生物量的测算. 水生生物学集刊, 1981, 47(3): 397-408.] |
| [19] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991.
|
| [20] |
王家辑. 中国淡水轮虫志. 北京: 科学出版社, 1961.
|
| [21] |
蒋燮治, 堵南山. 中国动物志——节肢动物门甲壳纲淡水枝角类. 北京: 科学出版社, 1979.
|
| [22] |
沈嘉瑞. 中国动物志——节肢动物门甲壳纲淡水桡足类. 北京: 科学出版社, 1979.
|
| [23] |
胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态. 北京: 科学出版社, 2006.
|
| [24] |
Qian KM, Liu X, Chen YW. A review on methods of cell enumeration and quantification of freshwater phytoplankton. J Lake Sci, 2015, 27(5): 767-775. [钱奎梅, 刘霞, 陈宇炜. 淡水浮游植物计数与定量方法. 湖泊科学, 2015, 27(5): 767-775. DOI:10.18307/2015.0502] |
| [25] |
张运林, 施坤, 秦伯强等. 一种大型浅水湖泊扰动强度空间格局的确定方法. CN105787263A, 2016-07-20.
|
| [26] |
Dai P, Liu K, Zhou YF et al. Zooplankton community characteristics in the lakeside zone of Wuli Lake, Taihu Lake. Journal of Hydroecology, 2019, 40(1): 55-63. [代培, 刘凯, 周彦锋等. 太湖五里湖湖滨带浮游动物群落结构特征. 水生态学杂志, 2019, 40(1): 55-63. DOI:10.15928/j.1674-3075.2019.01.009] |
| [27] |
Cai YJ, Xu H, Vilmi A et al. Relative roles of spatial processes, natural factors and anthropogenic stressors in structuring a lake macroinvertebrate metacommunity. Science of the Total Environment, 2017, 601/602: 1702-1711. DOI:10.1016/j.scitotenv.2017.05.264 |
| [28] |
Yang WH, Du X, Yao Z et al. Evaluation of the influencing factors of wuliangsuhai eutrophication based on VAR model. Environmental Science & Technology, 2021, 44(2): 25-32. [杨文焕, 杜璇, 姚植等. 基于VAR模型的乌梁素海富营养化影响因子评价. 环境科学与技术, 2021, 44(2): 25-32.] |
| [29] |
Du X, Wang QD, Zhang CW et al. Community structure of rotifers in relation to environmental factors in Lake Hongze. J Lake Sci, 2014, 26(2): 269-276. [都雪, 王齐东, 张超文等. 洪泽湖轮虫群落结构及其与环境因子的关系. 湖泊科学, 2014, 26(2): 269-276. DOI:10.18307/2014.0214] |
| [30] |
Chen Y, Peng K, Zhang QJ et al. Spatio-temporal distribution characteristics and driving factors of zooplankton in Hongze Lake. Environmental Science, 2021, 42(8): 3753-3762. [陈业, 彭凯, 张庆吉等. 洪泽湖浮游动物时空分布特征及其驱动因素. 环境科学, 2021, 42(8): 3753-3762. DOI:10.13227/j.hjkx.202010195] |
| [31] |
Suikkanen S, Laamanen M, Huttunen M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuarine, Coastal and Shelf Science, 2007, 71(3/4): 580-592. DOI:10.1016/j.ecss.2006.09.004 |
| [32] |
Li Q, Tian H, Jiang M et al. Community structure of zooplankton and influencing factors in Dianshan Lake of Shanghai. Journal of Hydroecology, 2015, 36(4): 69-77. [李强, 田华, 姜民等. 淀山湖浮游动物群落结构特征及其影响因子. 水生态学杂志, 2015, 36(4): 69-77. DOI:10.15928/j.1674-3075.2015.04.010] |
| [33] |
Wen CN, Huang W, Chen KN et al. Canonical correspondence analysis between zooplankton community structure and environmental factors in the littoral zone of Taihu Lake. Journal of Hydroecology, 2020, 41(2): 36-44. [温超男, 黄蔚, 陈开宁等. 太湖滨岸带浮游动物群落结构特征与环境因子的典范对应分析. 水生态学杂志, 2020, 41(2): 36-44. DOI:10.15928/j.1674-3075.2020.02.006] |
| [34] |
Wang YL, Yuan DN, Yuan GQ et al. Species diversity and community structure of zooplankton in summer and winter in the east lake, Wuhan. Acta Hydrobiologica Sinica, 2020, 44(4): 877-894. [王雨路, 袁丹妮, 袁国庆等. 武汉东湖夏冬两季浮游动物物种多样性及群落结构研究. 水生生物学报, 2020, 44(4): 877-894. DOI:10.7541/2020.105] |
| [35] |
Lü Q, Hu XR, Nie X et al. Impact of water level fluctuations on the succession of zooplankton in Poyang Lake. Acta Ecologica Sinica, 2020, 40(4): 1486-1495. [吕乾, 胡旭仁, 聂雪等. 鄱阳湖丰水期水位波动对浮游动物群落演替的影响. 生态学报, 2020, 40(4): 1486-1495.] |
| [36] |
Gao Y, Lai ZN, Li J et al. Spatial pattern of zooplankton diversity in Lianjiang River, Guangdong Province, China. Biodiversity Science, 2013, 21(6): 699-708. [高原, 赖子尼, 李捷等. 连江浮游动物多样性的空间分布. 生物多样性, 2013, 21(6): 699-708.] |
| [37] |
Chen JQ, Zhao K, Cao Y et al. Zooplankton community structure and its relationship with environmental factors in Poyang Lake. Acta Ecologica Sinica, 2020, 40(18): 6644-6658. [陈佳琪, 赵坤, 曹玥等. 鄱阳湖浮游动物群落结构及其与环境因子的关系. 生态学报, 2020, 40(18): 6644-6658.] |
| [38] |
Scheda SM, Cowell BC. Rotifer grazers and phytoplankton: Seasonal experiments on natural communities. Archiv fur Hydrobiologie, 1988, 114: 31-44. |
| [39] |
Mei HP, Wang ZL, Liu M et al. Characteristic water levels of Hongze Lake in the past five decades: Variation rules and influencing factors. Journal of Yangtze River Scientific Research Institute, 2021, 38(1): 35-40. [梅海鹏, 王振龙, 刘猛等. 洪泽湖近50a特征水位变化规律及影响因素. 长江科学院院报, 2021, 38(1): 35-40. DOI:10.11988/ckyyb.20191272] |
| [40] |
Liu BG, Wu JY, Hu Y et al. Seven years study of the seasonal dynamics of zooplankton communities in a large subtropical floodplain ecosystem: A Test of the PEG Model. Int J Environ Res Public Health, 2022, 19(2): 956. DOI:10.3390/ijerph19020956 |
| [41] |
Wu L, Feng WS, Chen XJ et al. Community structure characteristics of zooplankton in Yili region of Xinjiang in summer. Chinese Journal of Applied Ecology, 2008, 19(1): 163-172. [吴利, 冯伟松, 陈小娟等. 新疆伊犁地区夏季浮游动物群落结构特征. 应用生态学报, 2008, 19(1): 163-172.] |
| [42] |
Qiu XC, Zhao HX, Sun XX. Relationships between zooplankton and water environmental factors in Shahu Lake, Ningxia of Northwest China: A multivariate analysis. Chinese Journal of Ecology, 2012, 31(4): 896-901. [邱小琮, 赵红雪, 孙晓雪. 沙湖浮游动物与水环境因子关系的多元分析. 生态学杂志, 2012, 31(4): 896-901. DOI:10.13292/j.1000-4890.2012.0113] |
| [43] |
Cui C, Dong ZC, Tong J et al. Water quality evaluation and variation analysis of main channels of Hongze Lake. Jiangsu Water Resources, 2021(9): 27-34, 49. [崔璨, 董增川, 童建等. 洪泽湖主要入湖河道水质评价及变化规律. 江苏水利, 2021(9): 27-34, 49.] |
| [44] |
Cyr H, Sprules WG. The wind-driven distribution of nearshore zooplankton in a stratified lake varies with their body size. Freshwater Biology, 2022, 67(6): 991-1004. DOI:10.1111/fwb.13896 |
| [45] |
Sprules WG, Cyr H, Menza CW. Multiscale effects of wind-induced hydrodynamics on lake plankton distribution. Limnology and Oceanography, 2022, 67(7): 1631-1646. DOI:10.1002/lno.12158 |
| [46] |
Xue QJ, Rediske RR, Gong ZJ et al. Spatio-temporal variation of microcystins and its relationship to biotic and abiotic factors in Hongze Lake, China. Journal of Great Lakes Research, 2018, 44(2): 253-262. DOI:10.1016/j.jglr.2017.12.004 |
| [47] |
Fulton RS, Paerl HW. Toxic and inhibitory effects of the blue-green alga Microcystis aeruginosa on herbivorous zooplankton. Journal of Plankton Research, 1987, 9(5): 837-855. DOI:10.1093/plankt/9.5.837 |
| [48] |
Liu H, Xie P, Chen FZ et al. Enhancement of planktonic rotifers by Microcystis aeruginosa blooms: An enclosure experiment in a shallow eutrophic lake. Journal of Freshwater Ecology, 2002, 17(2): 239-247. DOI:10.1080/02705060.2002.9663892 |
| [49] |
Smith A, Gilbert KK. Relative susceptibilities of rotifers and cladocerans to Microcystis aeruginosa. Archiv fur Hydrobiologie, 1995, 132(3): 309-336. DOI:10.1127/archiv-hydrobiol/132/1995/309 |
| [50] |
耿红. 水体富营养化和蓝藻对轮虫影响的生态毒理学研究[学位论文]. 武汉: 中国科学院水生生物研究所, 2006.
|
| [51] |
Nian YG, Song YW, Li YJ et al. Regime shift theory and ecological restoration discussion in eutrophic shallow lakes. Research of Environmental Sciences, 2006, 19(1): 67-70. [年跃刚, 宋英伟, 李英杰等. 富营养化浅水湖泊稳态转换理论与生态恢复探讨. 环境科学研究, 2006, 19(1): 67-70. DOI:10.13198/j.res.2006.01.69.nianyg.017] |
| [52] |
Duan HX, Mao ZG, Wang GX et al. Ecological effects on enclosure culture demolition of Lake Hongze. J Lake Sci, 2021, 33(3): 706-714. [段海昕, 毛志刚, 王国祥等. 洪泽湖养殖网围拆除生态效应. 湖泊科学, 2021, 33(3): 706-714. DOI:10.18307/2021.0307] |
| [53] |
Wang XM, Song XM. Investigation on relevance between zooplankton in GaoTang lake and water environmental factors in Huainan city. Journal of Beijing Institute of Education: Natural Science, 2007, 2(5): 19-23. [王晓明, 宋晓梅. 淮南市高塘湖浮游动物与水环境因子的相关性研究. 北京教育学院学报: 自然科学版, 2007, 2(5): 19-23.] |
| [54] |
Visser AW, Stips A. Turbulence and zooplankton production: Insights from PROVESS. Journal of Sea Research, 2002, 47(3/4): 317-329. DOI:10.1016/S1385-1101(02)00120-X |
| [55] |
Visser AW, Mariani P, Pigolotti S. Swimming in turbulence: Zooplankton fitness in terms of foraging efficiency and predation risk. Journal of Plankton Research, 2008, 31(2): 121-133. DOI:10.1093/plankt/fbn109 |